Dendrit
Новости, статьи, учебные материалы по медицине
ПОДПИСАТЬСЯ НА РАССЫЛКУ
Регуляция обмена веществ. Механизм передачи гормонального сигнала в клетку.
Тема: "МЕХАНИЗМ ПЕРЕДАЧИ ГОРМОНАЛЬНОГО СИГНАЛА В КЛЕТКУ. ГОРМОНЫ ГИПОТАЛАМУСА И ГИПОФИЗА".
1. Определение понятия "гормоны", классификация и общие биологические признаки гормонов.
2. Классификация гормонов по химической природе, примеры.
3. Механизмы действия дистантных и проникающих в клетку гормонов.
4. Посредники действия гормонов на обмен веществ - циклические нуклеотиды (цАМФ, цГМФ), ионы Са2+, инозитолтрифосфат, рецепторные белки цитозоля. Реакции синтеза и распада цАМФ.
5. Каскадные механизмы активации ферментов, как способ усиления гормонального сигнала. Роль протеинкиназ.
6. Иерархия гормональной системы. Принцип обратной связи в регуляции секреции гормонов.
7. Гормоны гипоталамуса и передней доли гипофиза: химическая природа, механизм действия, ткани и клетки-мишени, биологический эффект.
23.1. Определение понятия “гормоны” и их классификация по химической природе.
23.1.1. Выучите определение понятия: гормоны – биологически активные соединения, выделяемые железами внутренней секреции в кровь или лимфу и оказывающие влияние на метаболизм клетки.
23.1.2. Запомните основные особенности действия гормонов на органы и ткани:
- гормоны синтезируются и выделяются в кровь специализированными эндокринными клетками;
- гормоны обладают высокой биологической активностью - физиологическое действие проявляется при концентрации их в крови порядка 10-6 - 10-12 моль/л;
- каждый гормон характеризуется присущей только ему структурой, местом синтеза и функцией; дефицит одного гормона не может быть восполнен другими веществами;
- гормоны, как правило, влияют на отдалённые от места их синтеза органы и ткани.
23.1.3. Гормоны осуществляют своё биологическое действие, образуя комплекс со специфическими молекулами – рецепторами. Клетки, содержащие рецепторы к определённому гормону, называются клетками-мишенями для этого гормона. Большинство гормонов взаимодействуют с рецепторами, расположенными на плазматической мембране клеток-мишеней; другие гормоны взаимодействуют с рецепторами, локализованными в цитоплазме и ядре клеток-мишеней. Имейте в виду, что дефицит как гормонов, так и их рецепторов может приводить к развитию заболеваний.
23.1.4. Некоторые гормоны могут синтезироваться эндокринными клетками в виде неактивных предшественников – прогормонов. Прогормоны могут запасаться в большом количестве в специальных секреторных гранулах и быстро активироваться в ответ на соответствующий сигнал.
23.1.5. Классификация гормонов основана на их химическом строении. Различные химические группы гормонов приведены в таблице 23.1.
| Класс химических веществ | Гормон или группа гормонов | Основное место синтеза |
|---|---|---|
| Белки и пептиды | Либерины Статины |
Гипоталамус |
| Вазопрессин Окситоцин |
Гипоталамус* | |
|
Тропные гормоны |
Передняя доля гипофиза (аденогипофиз) |
|
| Инсулин Глюкагон |
Поджелудочная железа (островки Лангерганса) | |
| Паратгормон | Паращитовидные железы | |
| Кальцитонин | Щитовидная железа | |
| Производные аминокислот | Иодтиронины (тироксин, трииодтиронин) |
Щитовидная железа |
| Катехоламины (адреналин, норадреналин) |
Мозговой слой надпочечников, симпатическая нервная система | |
| Стероиды | Глюкокортикоиды (кортизол) |
Кора надпочечников |
| Минералокортикоиды (альдостерон) |
Кора надпочечников | |
| Андрогены (тестостерон) |
Семенники | |
| Эстрогены (эстрадиол) |
Яичники | |
| Прогестины (прогестерон) |
Яичники |
* Местом секреции этих гормонов является задняя доля гипофиза (нейрогипофиз).
Следует иметь в виду, что кроме истинных гормонов выделяют также гормоны местного действия. Эти вещества синтезируются, как правило, неспециализированными клетками и оказывают свой эффект в непосредственной близости от места выработки (не переносятся током крови к другим органам). Примерами гормонов местного действия являются простагландины, кинины, гистамин, серотонин.
23.2. Иерархия регуляторных систем в организме.
23.2.1. Запомните, что в организме существует несколько уровней регуляции гомеостаза, которые тесно взаимосвязаны и функционируют как единая система (см. рисунок 23.1).
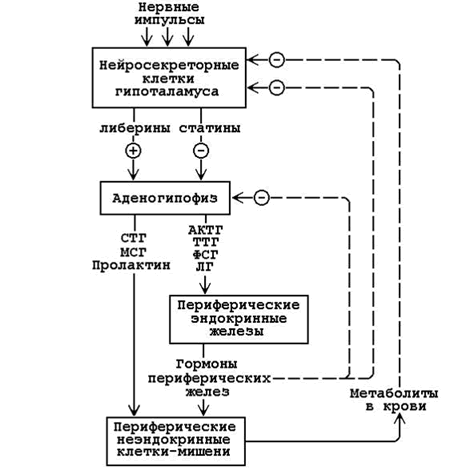
Рисунок 23.1. Иерархия регуляторных систем организма (пояснения в тексте).
23.2.2. 1. Сигналы из внешней и внутренней среды поступают в центральную нервную систему (высший уровень регуляции, осуществляет контроль в пределах целого организма). Эти сигналы трансформируются в нервные импульсы, попадающие на нейросекреторные клетки гипоталамуса. В гипоталамусе образуются:
- либерины (или рилизинг-факторы), стимулирующие секрецию гормонов гипофиза;
- статины – вещества, угнетающие секрецию этих гормонов.
Либерины и статины по системе портальных капилляров достигают гипофиза, где вырабатываются тропные гормоны. Тропные гормоны действуют на периферические ткани-мишени и стимулируют(знак “+”) образование и секрецию гормонов периферических эндокринных желёз. Гормоны периферических желёз угнетают (знак “–”) образование тропных гормонов, действуя на клетки гипофиза или нейросекреторные клетки гипоталамуса. Кроме того, гормоны, действуя на обмен веществ в тканях, вызывают изменения содержания метаболитов в крови, а те, в свою очередь, влияют (по механизму обратной связи) на секрецию гормонов в периферических железах (или непосредственно, или через гипофиз и гипоталамус).
2. Гипоталамус, гипофиз и периферические железы образуют средний уровень регуляции гомеостаза, обеспечивающий контроль нескольких метаболических путей в пределах одного органа, или ткани, или разных органов.
Гормоны эндокринных желёз могут влиять на обмен веществ:
- путём изменения количества ферментного белка;
- путём химической модификации ферментного белка с изменением его активности, а также
- путём изменения скорости транспорта веществ через биологические мембраны.
3. Внутриклеточные механизмы регуляции представляют собой низший уровень регуляции. Сигналами для изменения состояния клетки служат вещества, образующиеся в самих клетках или поступающие в неё.
23.3. Механизмы действия гормонов.
29.3.1. Обратите внимание, что механизм действия гормонов зависит от его химической природы и свойств – растворимости в воде или жирах. По механизму действия гормоны могут быть разделены на две группы: прямого и дистантного действия.
29.3.2. Гормоны прямого действия. К этой группе относятся липофильные (растворимые в жирах) гормоны – стероиды и йодтиронины. Эти вещества мало растворимы в воде и поэтому образуют в крови комплексные соединения с белками плазмы. К этим белкам относятся как специфические транспортные протеины (например, транскортин, связывающий гормоны коры надпочечников), так и неспецифические (альбумины).
Гормоны прямого действия в силу своей липофильности способны диффундировать через двойной липидный слой мембран клеток-мишеней. Рецепторы к этим гормонам находятся в цитозоле. Образующийсякомплекс гормона с рецептором перемещается в ядро клетки, где связывается с хроматином и воздействует на ДНК. В результате изменяется скорость синтеза РНК на матрице ДНК (транскрипция) и скорость образования специфических ферментативных белков на матрице РНК (трансляция). Это приводит к изменению количества ферментативных белков в клетках-мишенях и изменению в них направленности химических реакций (см. рисунок 2).
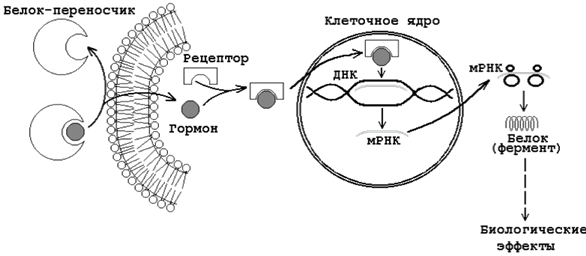
Рисунок 23.2. Механизм влияния на клетку гормонов прямого действия.
Как вам уже известно, регуляция синтеза белка может осуществляться при помощи механизмов индукции и репрессии.
Индукция синтеза белка происходит в результате стимуляции синтеза соответствующей матричной РНК. При этом возрастает концентрация определённого белка-фермента в клетке и увеличивается скорость катализируемых им химических реакций.
Репрессия синтеза белка происходит путём подавления синтеза соответствующей матричной РНК. В результате репрессии избирательно снижается концентрация определённого белка-фермента в клетке и уменьшается скорость катализируемых им химических реакций. Имейте в виду, что один и тот же гормон может вызывать индукцию синтеза одних белков и репрессию синтеза других белков. Эффект гормонов прямого действия обычно проявляется только спустя 2 - 3 часа после проникновения в клетку.
23.3.3. Гормоны дистантного действия. К гормонам дистантного действия относятся гидрофильные (растворимые в воде) гормоны – катехоламины и гормоны белково-пептидной природы. Так как эти вещества не растворимы в липидах, они не могут проникать через клеточные мембраны. Рецепторы для этих гормонов расположены на наружной поверхности плазматической мембраны клеток-мишеней. Гормоны дистантного действия реализуют своё действие на клетку при помощи вторичного посредника, в качестве которого чаще всего выступает циклический АМФ (цАМФ).
Циклический АМФ синтезируется из АТФ под действием аденилатциклазы:
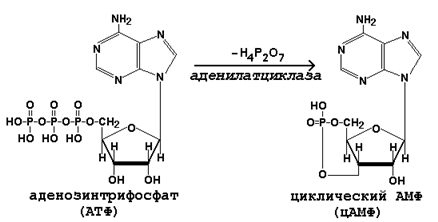
Механизм дистантного действия гормонов показан на рисунке 23.3.
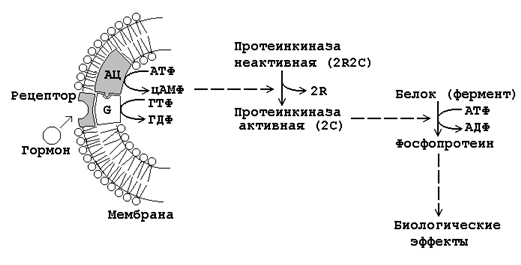
Рисунок 23.3. Механизм влияния на клетку гормонов дистантного действия.
Взаимодействие гормона с его специфическим рецептором приводит к активации G-белка клеточной мембраны. G-белок связывает ГТФ и активирует аденилатциклазу.
Активная аденилатциклаза превращает АТФ в цАМФ, цАМФ активирует протеинкиназу.
Неактивная протеинкиназа представляет собой тетрамер, который состоит из двух регуляторных (R) и двух каталитических (C) субъединиц. В результате взаимодействия с цАМФ происходит диссоциация тетрамера и освобождается активный центр фермента.
Протеинкиназа фосфорилирует белки-ферменты за счёт АТФ, либо активируя их, либо инактивируя. В результате этого изменяется (в одних случаях – увеличивается, в других – уменьшается) скорость химических реакций в клетках-мишенях.
Инактивация цАМФ происходит при участии фермента фосфодиэстеразы:
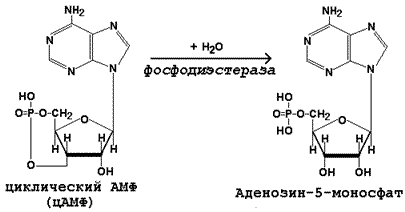
23.4. Гормоны гипоталамуса и гипофиза.
Как уже упоминалось, местом непосредственного взаимодействия высших отделов центральной нервной системы и эндокринной системы является гипоталамус. Это небольшой участок переднего мозга, который расположен непосредственно над гипофизом и связан с ним при помощи системы кровеносных сосудов, образующих портальную систему.
23.4.1. Гормоны гипоталамуса. В настоящее время известно, что нейросекреторные клетки гипоталамуса продуцируют 7 либеринов (соматолиберин, кортиколиберин, тиреолиберин, люлиберин, фоллиберин, пролактолиберин, меланолиберин) и 3 статина (соматостатин, пролактостатин, меланостатин). Все эти соединения являются пептидами.
Гормоны гипоталамуса через специальную портальную систему сосудов попадают в переднюю долю гипофиза (аденогипофиз). Либерины стимулируют, а статины подавляют синтез и секрецию тропных гормонов гипофиза. Эффект либеринов и статинов на клетки гипофиза опосредуется цАМФ- и Са2+-зависимыми механизмами.
Характеристика наиболее изученных либеринов и статинов приведена в таблице 23.2.
| Фактор | Место действия | Основные биологические эффекты | Регуляция секреции |
|---|---|---|---|
| Кортиколиберин | Аденогипофиз | Стимулирует секрецию адренокортикотропного гормона (АКТГ) | Секреция стимулируется при стрессах и подавляется АКТГ |
| Тиреолиберин | - “ – “ - | Стимулирует секрецию тиреотропного гормона (ТТГ) и пролактина | Секрецию тормозят тиреоидные гормоны |
| Соматолиберин | - “ – “ - | Стимулирует секрецию соматотропного гормона (СТГ) | Секрецию стимулирует гипогликемия |
| Люлиберин | - “ – “ - | Стимулирует секрецию фолликулостимулирующего гормона (ФСГ) и лютеинизирующего гормона (ЛГ) | У мужчин секреция вызывается снижением содержания тестостерона в крови, у женщин – снижением концентрации эстрогенов. Высокая концентрация ЛГ и ФСГ в крови подавляет секрецию |
| Соматостатин | - “ – “ - | Тормозит секрецию СТГ и ТТГ | Секреция вызывается физической нагрузкой. Фактор быстро инактивируется в тканях тела. |
| Пролактостатин | - “ – “ - | Тормозит секрецию пролактина | Секрецию стимулирует высокая концентрация пролактина и подавляют эстрогены, тестостерон и нервные сигналы при сосании. |
| Меланостатин | - “ – “ - | Угнетает секрецию МСГ (меланоцитостимулирующего гормона) | Секрецию стимулирует меланотонин |
23.4.2. Гормоны аденогипофиза. Аденогипофиз (передняя доля гипофиза) продуцирует и выделяет в кровь ряд тропных гормонов, регулирующих функцию как эндокринных, так и неэндокринных органов. Все гормоны гипофиза являются белками или пептидами. Внутриклеточным посредником всех гипофизарных гормонов (кроме соматотропина и пролактина) служит циклический АМФ (цАМФ). Характеристика гормонов передней доли гипофиза приводится в таблице 3.
| Гормон | Ткань-мишень | Основные биологические эффекты | Регуляция секреции |
|---|---|---|---|
| Адренокортикотропный гормон (АКТГ) | Кора надпочечников | Стимулирует синтез и секрецию стероидов корой надпочечников | Стимулируется кортиколиберином |
| Тиреотропный гормон (ТТГ) | Щитовидная железа | Усиливает синтез и секрецию тиреоидных гормонов | Стимулируется тиреолиберином и подавляется тиреоидными гормонами |
| Соматотропный гормон (гормон роста, СТГ) | Все ткани | Стимулирует синтез РНК и белка, рост тканей, транспорт глюкозы и аминокислот в клетки, липолиз | Стимулируется соматолиберином, подавляется соматостатином |
| Фолликулостимулирующий гормон (ФСГ) | Семенные канальцы у мужчин, фолликулы яичников у женщин | У мужчин повышает образование спермы, у женщин – образование фолликулов | Стимулируется люлиберином |
| Лютеинизирующий гормон (ЛГ) | Интерстициальные клетки семенников (у мужчин) и яичников (у женщин) | Вызывает секрецию эстрогенов, прогестерона у женщин, усиливает синтез и секрецию андрогенов у мужчин | Стимулируется люлиберином |
| Пролактин | Молочные железы (альвеолярные клетки) | Стимулирует синтез белков молока и развитие молочных желёз | Подавляется пролактостатином |
| Меланоцитостимулирующий гормон (МСГ) | Пигментные клетки | Повышает синтез меланина в меланоцитах (вызывает потемнение кожи) | Подавляется меланостатином |
23.4.3. Гормоны нейрогипофиза. К гормонам, секретируемым в кровоток задней долей гипофиза, относятся окситоцин и вазопрессин. Оба гормона синтезируются в гипоталамусе в виде белков-предшественников и перемещаются по нервным волокнам в заднюю долю гипофиза.
Окситоцин – нонапептид, вызывающий сокращения гладкой мускулатуры матки. Он используется в акушерстве для стимуляции родовой деятельности и лактации.
Вазопрессин – нонапептид, выделяемый в ответ на повышение осмотического давления крови. Клетками-мишенями для вазопрессина являются клетки почечных канальцев и гладкомышечные клетки сосудов. Действие гормона опосредовано цАМФ. Вазопрессин вызывает сужение сосудов и повышение артериального давления, а также усиливает реабсорбцию воды в почечных канальцах, что приводит к снижению диуреза.
23.4.4. Основные виды нарушений гормональной функции гипофиза и гипоталамуса. При дефиците соматотропного гормона, возникающем в детском возрасте, развивается карликовость (низкий рост). При избытке соматотропного гормона, возникающем в детском возрасте, развивается гигантизм (аномально высокий рост).
При избытке соматотропного гормона, возникающем у взрослых (в результате опухоли гипофиза), развивается акромегалия – усиленный рост кистей рук, ступней, нижней челюсти, носа.
При недостатке вазопрессина, возникающем вследствие нейротропных инфекций, черепно-мозговых травм, опухолей гипоталамуса, развивается несахарный диабет. Основным симптомом этого заболевания является полиурия – резкое увеличение диуреза при пониженной (1,001 – 1,005) относительной плотности мочи.
28.4. Гормоны поджелудочной железы.
Обратите внимание, что эндокринная часть поджелудочной железы продуцирует и выделяет в кровь гормоны инсулин и глюкагон.
1. Инсулин. Инсулин – белково-пептидный гормон, вырабатываемый β-клетками островков Лангерганса. Молекула инсулина состоит из двух полипептидных цепей (А и В), содержащих 21 и 30 аминокислотных остатков соответственно; цепи инсулина связаны между собой двумя дисульфидными мостиками. Образуется инсулин из белка-предшественника (препроинсулина) путём частичного протеолиза (см. рисунок 4). После отщепления сигнальной последовательности образуется проинсулин. В результате ферментативного превращения удаляется фрагмент полипептидной цепи, содержащий около 30 аминокислотных остатков (С-пептид), и образуется инсулин.
Стимулом для секреции инсулина является гипергликемия – повышение содержания глюкозы в крови (например, после приёма пищи). Главные мишени для инсулина – клетки печени, мышц и жировой ткани. Механизм действия – дистантный.
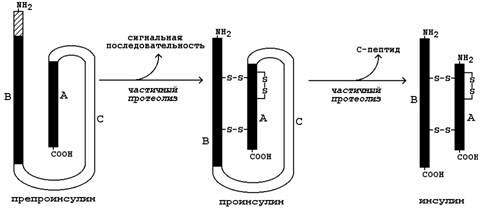
Рисунок 4. Схема превращения препроинсулина в инсулин.
Рецептор инсулина представляет собой сложный белок – гликопротеин, расположенный на поверхности клетки-мишени. Этот белок состоит их двух α-субъединиц и двух β-субъединиц, связанных между собой дисульфидными мостиками. β-Субъединицы содержат несколько аминокислотных остатков тирозина. Рецептор инсулина обладает тирозинкиназной активностью, т.е. способен катализировать перенос остатков фосфорной кислоты от АТФ на ОН-группу тирозина (рисунок 5).
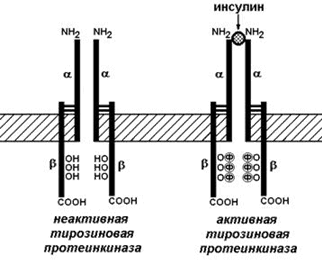
Рисунок 5. Инсулиновый рецептор.
В отсутствие инсулина рецептор не проявляет ферментативной активности. При связывании с инсулином рецептор подвергается аутофосфорилированию, т.е. β-субъединицы фосфорилируют друг друга. В результате изменяется конформация рецептора и он приобретает способность фосфорилировать другие внутриклеточные белки. В дальнейшем комплекс инсулина с рецептором погружается в цитоплазму и его компоненты расщепляются в лизосомах.
Образование гормон-рецепторного комплекса повышает проницаемость клеточных мембран для глюкозы и аминокислот. Под действием инсулина в клетках-мишенях:
а) снижается активность аденилатциклазы и увеличивается активность фосфодиэстеразы, что приводит к понижению концентрации цАМФ;
б) повышается скорость окисления глюкозы и снижается скорость глюконеогенеза;
в) увеличивается синтез гликогена и жиров и подавляется их мобилизация;
г) ускоряется синтез белка и тормозится его распад.
Все эти изменения направлены на ускоренное использование глюкозы, что приводит к снижению содержания глюкозы в крови. Инактивация инсулина происходит главным образом в печени и заключается в разрыве дисульфидных связей между цепями А и В.
2. Глюкагон. Глюкагон – полипептид, содержащий 29 аминокислотных остатков. Он продуцируется α-клетками островков Лангерганса в виде белка-предшественнника (проглюкагона). Частичный протеолиз прогормона и секреция глюкагона в кровь происходит при гипогликемии, вызванной голоданием.
Клетки-мишени для глюкагона – печень, жировая ткань, миокард. Механизм действия – дистантный (посредником является цАМФ).
Под действием глюкагона в клетках-мишенях:
а) ускоряется мобилизация гликогена в печени (см. рисунок 6) и тормозится его синтез;
б) ускоряется мобилизация жиров (липолиз) в жировой ткани и тормозится их синтез;
в) угнетается синтез белка и усиливается его катаболизм;
г) ускоряется глюконеогенез и кетогенез в печени.
Конечный эффект глюкагона – поддержание высокого уровня глюкозы в крови.
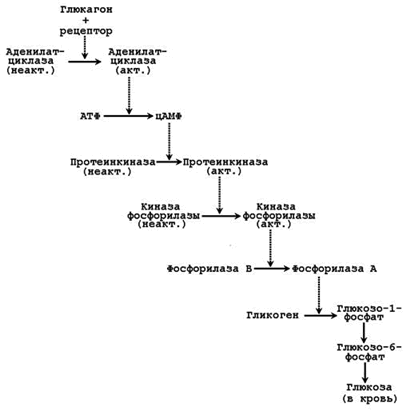
Рисунок 6. Каскадный механизм активации фосфорилазы гликогена под влиянием глюкагона.
3. Нарушения гормональной функции поджелудочной железы. Наиболее часто встречается сахарный диабет – заболевание, обусловленное нарушением синтеза и секреции инсулина β-клетками (диабет I типа) либо дефицитом инсулинчувствительных рецепторов в клетках-мишенях (диабет II типа). Для сахарного диабета характерны следующие нарушения обмена веществ:
а) снижение использования глюкозы клетками, усиление мобилизации гликогена и активация глюконеогенеза в печени приводят к увеличению содержания глюкозы в крови (гипергликемия) и преодоление ею почечного порога (глюкозурия);
б) ускорение липолиза (расщепления жиров), избыточное образование ацетил-КоА, используемого для синтеза с последующим поступлением в кровь холестерола (гиперхолестеролемия) и кетоновых тел (гиперкетонемия); кетоновые тела легко проникают в мочу (кетонурия);
в) снижение скорости синтеза белка и усиление катаболизма аминокислот в тканях приводит к повышению концентрации мочевины и других азотистых веществ в крови (азотемия) и увеличению их выведения с мочой (азотурия);
г) выведение почками больших количеств глюкозы, кетоновых тел и мочевины сопровождается увеличением диуреза (полиурия).
28.5. Гормоны мозгового вещества надпочечников.
К гормонам мозгового вещества надпочечников относятся адреналин и норадреналин (катехоламины). Они синтезируются в хромаффинных клетках из тирозина (рисунок 7).
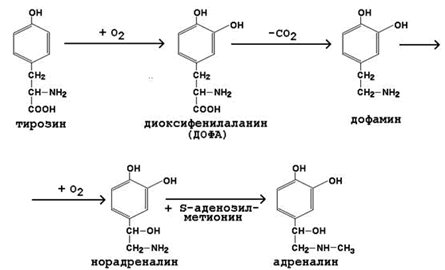
Рисунок 7. Схема синтеза катехоламинов.
Секреция адреналина усиливается при стрессе, физических нагрузках. Мишени для катехоламинов – клетки печени, мышечной и жировой ткани, сердечно-сосудистая система. Механизм действия – дистантный. Эффекты реализуются через аденилатциклазную систему и проявляются изменениями углеводного обмена. Подобно глюкагону, адреналин вызывает активацию мобилизации гликогена (см. рисунок 6) в мышцах и печени, липолиз в жировой ткани. Это приводит к увеличению содержания глюкозы, лактата и жирных кислот в крови. Адреналин усиливает также сердечную деятельность, вызывает сужение сосудов.
Обезвреживание адреналина происходит в печени. Основными путями обезвреживания являются: метилирование (фермент – катехол-орто-метилтрансфераза, КОМТ), окислительное дезаминирование (фермент – моноаминооксидаза, МАО) и конъюгация с глюкуроновой кислотой. Продукты обезвреживания выводятся с мочой.
ОБНОВЛЕНИЯ
08 июня 2019, 16:18:59
08 июня 2019, 16:10:24
07 июня 2019, 22:59:05
09 мая 2019, 16:13:23
27 апреля 2019, 16:00:00
24 апреля 2019, 01:02:18
17 апреля 2019, 22:26:23
17 апреля 2019, 22:21:37
17 апреля 2019, 22:07:42
17 апреля 2019, 22:06:28
ПОДПИСАТЬСЯ НА РАССЫЛКУ
ПРЕДМЕТЫ
- Анатомия
- Акушерство и гинекология
- БЖД, медицина катастроф
- Биохимия
- Биология
- Гистология
- Гигиена
- Генетика
- Диетология
- Дерматовенерология
- Инфекционные болезни
- Культурология
- Лабораторная диагностика
- Летняя практика
- Лучевая диагностика
- Медицинская информатика
- Микробиология
- Неврология
- Общественное здоровье
- Общий уход
О НАС
«Dendrit» - информационный портал для медицинских работников, студентов медицинских ВУЗов, исследователей и пациентов.
Ваш источник новостей и знаний о здоровье.
dendrit.ru © 2014-2024
