Dendrit
Новости, статьи, учебные материалы по медицине
ПОДПИСАТЬСЯ НА РАССЫЛКУ
Пентозофосфатный путь. Глюконеогенез. Биосинтез и мобилизация гликогена
Тема: «ПЕНТОЗОФОСФАТНЫЙ ПУТЬ. ГЛЮКОНЕОГЕНЕЗ. БИОСИНТЕЗ И МОБИЛИЗАЦИЯ ГЛИКОГЕНА»
1. Пентозофосфатный (апотомический) путь окисления глюкозо-6-фосфата: биологическая роль, локализация в клетке, основные этапы. Последовательность реакций окислительного этапа пентозофосфатного пути.
2. Глюконеогенез: биологическая роль, локализация в клетке и тканях, последовательность реакций, возможные предшественники, регуляция, баланс АТФ.
3. Биосинтез гликогена: биологическая роль, локализация в клетке и тканях, последовательность реакций, гормональная регуляция. УДФ-глюкоза – образование и использование в обмене углеводов.
4. Мобилизация гликогена: биологическая роль, локализация в тканях, последовательность реакций, гормональная регуляция.
| Раздел 16.1 |
Пентозофосфатный (апотомический) путь окисления глюкозы. |
|
16.1.1. Пентозофосфатный путь представляет собой прямое окисление глюкозы и протекает в цитоплазме клеток. Наибольшая активность ферментов пентозофосфатного пути обнаружена в клетках печени, жировой ткани, коры надпочечников, молочной железы в период лактации, зрелых эритроцитах. Низкий уровень этого процесса выявлен в скелетных и сердечной мышцах, мозге, щитовидной железе, легких. Пентозофосфатный путь называют также апотомическим путём, так как в его реакциях происходит укорочение углеродной цепи гексозы на один атом, который включается в молекулу СО2. 16.1.2. Пентозофосфатный путь выполняет в организме две важнейшие метаболические функции: он является главным источником НАДФН для синтеза жирных кислот, холестерола, стероидных гормонов, микросомального окисления; в эритроцитах НАДФН используется для восстановления глутатиона – вещества, препятствующего пероксидному гемолизу;
он является главным источником пентоз для синтеза нуклеотидов, нуклеиновых кислот, коферментов (АТФ, НАД, НАДФ, КоА-SН и др.).
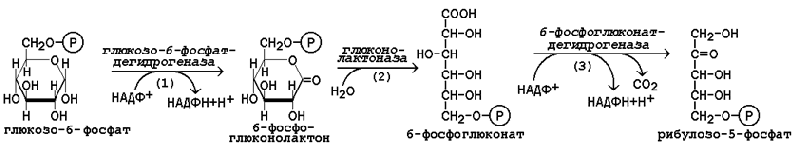
16.1.3. В пентозофосфатном пути можно выделить две фазы - окислительную и неокислительную. Исходным субстратом окислительной фазы является глюкозо-6-фосфат, который непосредственно подвергается дегидрированию с участием НАДФ-зависимой дегидрогеназы (рисунок 16.1, реакция 1). Продукт реакции гидролизуется (реакция 2), а образующийся 6-фосфоглюконат дегидрируется и декарбоксилируется (реакция 3). Таким образом, происходит укорочение углеродной цепи моносахарида на один углеродный атом («апотомия»), и образуется рибулозо-5-фосфат.
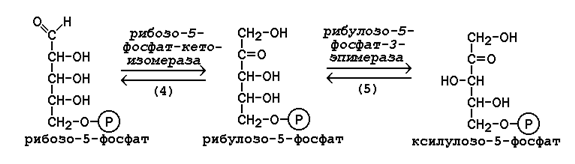
Рисунок 16.1. Реакции окислительной фазы пентозофосфатного пути. 16.1.4. Неокислительная фаза пентозофосфатного пути начинается с реакций изомеризации. В ходе этих реакций одна часть рибулозо-5-фосфата изомеризуется в рибозо-5-фосфат, другая - в ксилулозо-5-фосфат (рисунок 16.2, реакции 4 и 5).
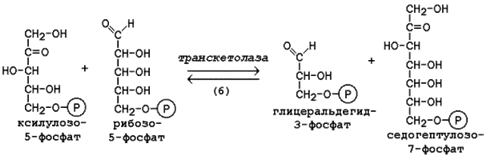
Следуюшая реакция протекает при участии фермента транскетолазы, коферментом которой является тиаминдифосфат (производное витамина B1). В этой реакции происходит перенос двухуглеродного фрагмента с ксилулозо-5-фосфата на рибозо-5-фосфат:
Образовавшиеся продукты взаимодействуют между собой в реакции, которая катализируется трансальдолазой и заключается а переносе остатка дигидроксиацетона на глицеральдегид-3-фосфат.
16.1.5. Посредством пентозофосфатного пути может происходить полное окисление глюкозо-6-фосфата до шести молекул СО2. Все эти молекулы образуются из С-1-атомов шести молекул глюкозо-6-фосфата, а из образовавшихся при этом шести молекул рибулозо-5-фосфата снова регенерируются пять молекул глюкозо-6-фосфата:
Если упростить представленную схему, то получится:
Таким образом, полное окисление 1 молекулы глюкозы в пентозофосфатном пути сопровождается восстановлением 12 молекул НАДФ. |
|

| Раздел 16.2 |
Глюконеогенез. |
|
|
|
|
16.2.1. Глюконеогенез - биосинтез глюкозы из различных соединений неуглеводной природы. Биологическая роль глюконеогенеза заключается в поддержании постоянного уровня глюкозы в крови, что необходимо для нормального энергообеспечения тканей, для которых характерна непрерывная потребность в углеводах. Особенно это касается центральной нервной системы. Роль глюконеогенеза возрастает при недостаточном поступлении углеводов с пищей. Так, в организме голодающего человека может синтезироваться до 200 г глюкозы в сутки. Глюконеогенез быстрее, чем другие метаболические процессы, реагирует на изменения диеты: введение с пищей большого количества белков и жиров активизирует процессы глюконеогенеза; избыток углеводов, наоборот, тормозит новообразование глюкозы. Интенсивные физические нагрузки сопровождаются быстрым истощением запасов глюкозы в организме. В этом случае глюконеогенез является основным путём пополнения углеводных ресурсов, предупреждая развитие гипогликемии. Глюконеогенез в организме тесно связан также с процессами обезвреживания аммиака и поддержанием кислотно-основного баланса. 16.2.2. Основным местом биосинтеза глюкозы de novo является печень. Глюконеогенез протекает также в корковом слое почек. Принято считать, что вклад почек в глюконеогенез в физиологических условиях составляет около 10% глюкозы, синтезируемой в организме; при патологических состояниях эта доля может значительно возрастать. Незначительная активность ферментов глюконеогенеза обнаружена в слизистой тонкого кишечника. 16.2.3. Последовательность реакций глюконеогенеза представляет собой обращение соответствующих реакций гликолиза. Лишь три реакции гликолиза необратимы вследствие происходящих в ходе их значительных энергетических сдвигов:
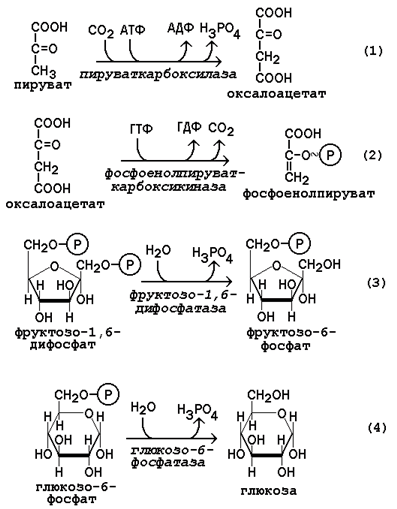
Обход этих энергетических барьеров обеспечивают ключевые ферменты глюконеогенеза. Обратный переход пирувата в фосфоенолпируват требует участия двух ферментов. Первый из них – пируваткарбоксилаза - катализирует реакцию образования оксалоацетата (рисунок 16.4, реакция 1). Коферментом пируваткарбоксилазы является биотин (витамин Н). Реакция протекает в митохондриях. Роль её заключается также в пополнении фонда оксалоацетата для цикла Кребса. Все последующие реакции глюконеогенеза протекают в цитоплазме. Мембрана митохондрий непроницаема для оксалоацетата, и он переносится в цитоплазму в виде других метаболитов: малата или аспартата. В цитоплазме указанные соединения вновь переходят в оксалоацетат. При участии фосфоенолпируваткарбоксикиназы из оксалоацетата образуется фосфоенолпируват (рисунок 16.4, реакция 2). Фосфоенолпируват в результате обращения ряда реакций гликолиза переходит во фруктозо-1,6-дифосфат. Превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат катализируетсяфруктозодифосфатазой (рисунок 16.4, реакция 3). Фруктозо-6-фосфат изомеризуется в глюкозо-6-фосфат. Заключительной реакцией глюконеогенеза является гидролиз глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатазы (рисунок 16.4, реакция 4).
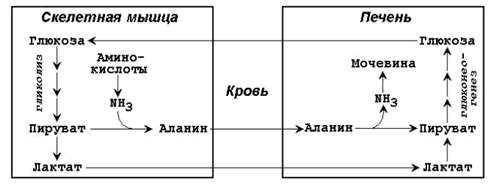
Рисунок 16.4. Обходные реакции глюконеогенеза. 16.2.4. Основными источниками глюкозы в глюконеогенезе являются лактат, аминокислоты, глицерол и метаболиты цикла Кребса. Лактат – конечный продукт анаэробного окисления глюкозы. Может включаться в глюконеогенез после окисления до пирувата в лактатдегидрогеназной реакции (см. раздел «Гликолиз», рисунок 15.4, реакция 11). При продолжительной физической работе основным источником лактата является скелетная мускулатура, в клетках которой преобладают анаэробные процессы. Накопление молочной кислоты в мышцах ограничивает их работоспособность. Это связано с тем, что при повышении концентрации молочной кислоты в ткани снижается уровень рН (молочнокислый ацидоз). Изменение рН приводит к ингибированию ферментов важнейших метаболических путей. В утилизации образующейся молочной кислоты важное место принадлежитглюкозо-лактатному циклу Кори (рисунок 16.5).
Рисунок 16.5. Цикл Кори и глюкозо-аланиновый цикл (пояснения в тексте). Глюкогенные аминокислоты, к которым относятся большинство белковых аминокислот. Ведущее место в глюконеогенезе среди аминокислот принадлежит аланину, который может превращаться в пируват путём трансаминирования. При голодании, физической работе и других состояниях в организме функционирует глюкозо-аланиновый цикл, подобный циклу Кори для лактата (рисунок 16.2). Существование цикла аланин – глюкоза препятствует отравлению организма, так как в мышцах нет ферментов, утилизирующих аммиак. В результате тренировки мощность этого цикла значительно возрастает. Другие аминокислоты могут, подобно аланину, превращаться в пируват, а также в промежуточные продукты цикла Кребса (α-кетоглутарат, фумарат, сукцинил-КоА). Все эти метаболиты способны преобразовываться в оксалоацетат и включаться в глюконеогенез. Глицерол – продукт гидролиза липидов в жировой ткани. Этот процесс значительно усиливается при голодании. В печени глицерол превращается в диоксиацетонфосфат – промежуточный продукт гликолиза и может быть использован в глюконеогенезе. Жирные кислоты и ацетил-КоА не являются предшественниками глюкозы. Окисление этих соединений обеспечивает энергией процесс синтеза глюкозы. 16.2.5. Энергетический баланс. Путь синтеза глюкозы из пирувата (рисунок 16.6) содержит три реакции, сопровождающиеся потреблением энергии АТФ или ГТФ:
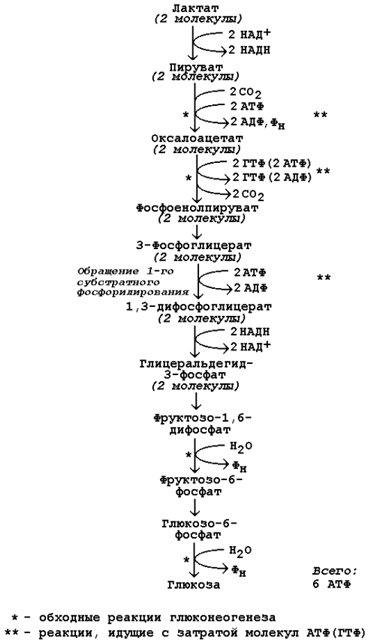
Каждая из этих реакций повторяется дважды, так как для образования 1 молекулы глюкозы (С6) используются 2 молекулы пирувата (С3). Поэтому энергетический баланс синтеза глюкозы из пирувата составляет – 6 молекул нуклеозидтрифосфатов (4 молекулы АТФ и 2 молекулы ГТФ). При использовании других предшественников энергетический баланс биосинтеза глюкозы отличается.
Рисунок 16.6. Энергетический баланс биосинтеза глюкозы из лактата. 16.2.6. Регуляция глюконеогенеза. Скорость глюконеогенеза определяется доступностью субстратов – предшественников глюкозы. Увеличение концентрации в крови любого из предшественников глюкозы приводит к стимуляции глюконеогенеза. Некоторые метаболиты являются аллостерическими эффекторами ферментов глюконеогенеза. Например, ацетил-КоА в повышенных концентрациях аллостерически активирует пируваткарбоксилазу, катализирующую первую реакцию глюконеогенеза. Аденозинмонофосфат, наоборот, оказывает ингибирующее действие на фруктозодифосфатазу, а избыток глюкозы ингибирует глюкозо-6-фосфатазу. Гормон поджелудочной железы глюкагон, гормоны надпочечников адреналин и кортизол повышают скорость биосинтеза глюкозы в организме, увеличивая активность ключевых ферментов глюконеогенеза либо увеличивая концентрацию этих ферментов в клетках. Гормон поджелудочной железы инсулин способствует снижению скорости глюконеогенеза в организме. |
|
| Раздел 16.3 |
Биосинтез гликогена. |
|
|
|
|
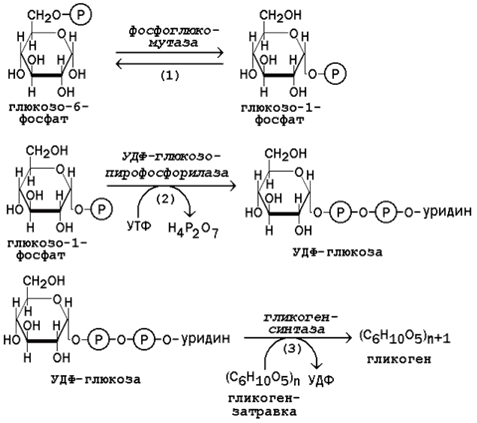
16.3.1. Гликоген – биополимер, состоящий из остатков глюкозы, он является компонентом всех тканей животных и человека (см. рисунок 15.1). Этот полисахарид служит основным источником энергии и резервом углеводов в организме. Содержание гликогена в различных органах зависит от физиологического состояния организма. Наиболее высокое содержание гликогена обнаруживается в печени (от 2 до 6% от массы органа). Хотя концентрация гликогена в мышцах значительно ниже (от 0,5 до 1,5%), однако в норме его количество в мышцах составляет 2/3 от общего его содержания в организме. Гликоген отличается значительным разнообразием по структуре и по относительной молекулярной массе. Молекулы его ветвисты. Глюкозные остатки образуют цепи, в которых они связаны между собой α-1,4-гликозидными связями. Разветвления образуются при помощи α-1,6-гликозидных связей. Большая часть глюкозы, поступающей в организм с пищей, превращается в клетках печени в гликоген. Это связано с тем, что накопление легко растворимой глюкозы в клетках привело бы к резкому увеличению осмотического давления и разрушению клеточной мембраны. 16.3.2. Молекула глюкозы фосфорилируется при участии фермента глюкокиназы (в гепатоцитах) или гексокиназы (в других клетках) с образованием глюкозо-6-фосфата. Этот метаболит под влиянием фосфоглюкомутазы превращается в глюкозо-1-фосфат (рисунок 16.7, реакция 1).
Рисунок 16.7. Реакции биосинтеза гликогена. Глюкозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ) в присутствии фермента УДФ-глюкозо-пирофосфорилазы с образованием УДФ-глюкозы – ключевого метаболита углеводного обмена (рисунок 16.7, реакция 2). Перенос глюкозного остатка с УДФ-глюкозы на затравочную полисахаридную цепь осуществляет гликогенсинтаза – регуляторный фермент данного метаболического пути (рисунок 16.7, реакция 3). Гликогенсинтаза существует в двух формах: гликогенсинтаза b (неактивная) и гликогенсинтаза а (активная). Превращение формы а в форму b катализируется цАМФ-зависимой протеинкиназой, обратный процесс – протеинфосфатазой. Образование α-1,6-гликозидных связей в молекуле гликогена катализирует ветвящий фермент или трансглюкозидаза (рисунок 16.8) .
Рисунок 16.8. Образование α-1,6-гликозидных связей в молекуле гликогена. |
|
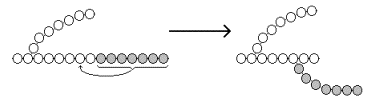
| Раздел 16.4 |
Мобилизация гликогена и гликогенолиз. |
|
|
|
|
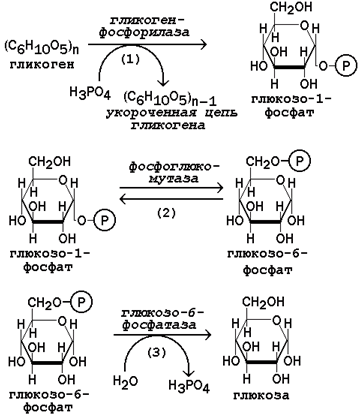
16.4.1. Мобилизация, или распад гликогена – процесс превращения гликогена в глюкозу, происходящий в печени. Таким образом, распад гликогена в печени наряду с глюконеогенезом принимает участие в поддержании уровня глюкозы в крови.
Рисунок 16.9. Реакции мобилизации гликогена. Первую реакцию внутриклеточного расщепления гликогена катализирует фермент фосфорилаза гликогена (рисунок 16.9, реакция 1). Простетической группой его является пиридоксальфосфат (производное витамина В6). Фосфорилаза гликогена – регуляторный фермент, он существует в двух формах: фосфорилаза а (активная) и фосфорилаза b (неактивная). Переход фосфорилазы b в фосфорилазу акатализирует киназа фосфорилазы, которая фосфорилирует неактивный фермент. Превращение фосфорилазы а в фосфорилазу b катализирует протеинфосфатаза, которая осуществляет дефосфорилирование активного фермента. Следует отметить, что ускорение процессов мобилизации гликогена происходит одновременно с торможением его биосинтеза, и наоборот. Активацию фосфорилазы гликогена вызывают гормоны адреналин и глюкагон. Фосфорилаза расщепляет в молекуле гликогена лишь α-1,4-гликозидные связи. Расщепление α-1,6-гликозидных связей осуществляется амило-1,6-гликозидазой гидролитически с образованием свободной глюкозы. Глюкозо-1-фосфат, образующийся в фосфорилазной реакции, при участии уже известного Вам фермента фосфоглюкомутазы переходит в глюкозо-6-фосфат (рисунок 16.9, реакция 2). 16.4.2. Дальнейшая судьба глюкозо-6-фосфата, образовавшегося при распаде гликогена, в разных тканях различна. Как уже упоминалось, в печени и почках имеется фермент глюкозо-6-фосфатаза, отщепляющая фосфат от глюкозо-6-фосфата (рисунок 16.9, реакция 3). Образовавшаяся глюкоза диффундирует в кровь, откуда поглощается клетками организма и служит для них источником энергии. В мышечной ткани фермент глюкозо-6-фосфатаза отсутствует. Поэтому глюкозо-6-фосфат, образовавшийся при распаде гликогена, подвергается дальнейшему превращению по гликолитическому пути с образованием лактата (этот процесс называется гликогенолизом). Образующийся в реакциях субстратного фосфорилирования АТФ используется в процессах мышечного сокращения. Таким образом, гликоген мышц является резервом энергии только для мышечной ткани. |
|
| С.М.Ершиков, 2007-2011. | |
ОБНОВЛЕНИЯ
08 июня 2019, 16:18:59
08 июня 2019, 16:10:24
07 июня 2019, 22:59:05
09 мая 2019, 16:13:23
27 апреля 2019, 16:00:00
24 апреля 2019, 01:02:18
17 апреля 2019, 22:26:23
17 апреля 2019, 22:21:37
17 апреля 2019, 22:07:42
17 апреля 2019, 22:06:28
ПОДПИСАТЬСЯ НА РАССЫЛКУ
ПРЕДМЕТЫ
- Анатомия
- Акушерство и гинекология
- БЖД, медицина катастроф
- Биохимия
- Биология
- Гистология
- Гигиена
- Генетика
- Диетология
- Дерматовенерология
- Инфекционные болезни
- Культурология
- Лабораторная диагностика
- Летняя практика
- Лучевая диагностика
- Медицинская информатика
- Микробиология
- Неврология
- Общественное здоровье
- Общий уход
О НАС
«Dendrit» - информационный портал для медицинских работников, студентов медицинских ВУЗов, исследователей и пациентов.
Ваш источник новостей и знаний о здоровье.
dendrit.ru © 2014-2024
