Dendrit
Новости, статьи, учебные материалы по медицине
ПОДПИСАТЬСЯ НА РАССЫЛКУ
Структурно-функциональная организация ферментов. Регуляция активности ферментов. Определение амилазной активности мочи.
Тема: "СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ФЕРМЕНТОВ. РЕГУЛЯЦИЯ АКТИВНОСТИ ФЕРМЕНТОВ. ОПРЕДЕЛЕНИЕ АМИЛАЗНОЙ АКТИВНОСТИ МОЧИ".
1. Понятие о простых и сложных ферментах. Состав сложных ферментов.
2. Коферменты. Классификация коферментов: витаминсодержащие и невитаминсо-держащие (пептидные и нуклеотидные), примеры. Функции коферментов.
3. Активный центр фермента. Функциональные группы, входящие в его состав. Контактный и каталитический участки активного центра. Типы химических взаимодействий при образовании фермент-субстратного комплекса.
4. Аллостерический (регуляторный) центр фермента, его функция. Роль третичной и четвертичной структуры молекулы белка-фермента в образовании активных и аллостерических центров.
5. Понятие об активаторах и ингибиторах ферментов. Роль ионов металлов в регуля-ции активности ферментов (примеры).
6. Виды ингибирования ферментов (обратимое и необратимое, конкурентное и неконкурентное). Примеры.
7. Основные механизмы регуляции активности ферментов: ковалентная модификация (фосфорилирование-дефосфорилирование, частичный протеолиз); кооперативные и аллостерические эффекты; регуляция по принципу отрицательной обратной связи.
8. Структурно-функциональная организация ферментов. Изоферменты: биологическая роль, структурная организация, использование изоферментов лактатдегидрогеназы в энзимодиагностике. Мультиферменты: биологическая роль, структурная организация (на примере пируватдегидрогеназного комплекса).
|
Раздел 8.1 |
Понятие о простых и сложных ферментах. Состав сложных ферментов. Коферменты: классификация и функции. |
||||||||||||
|
|
|||||||||||||
|
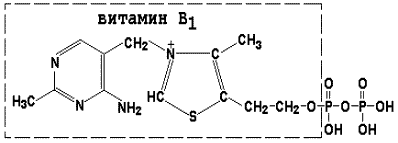
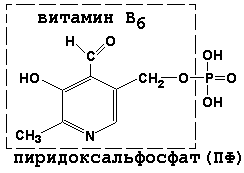
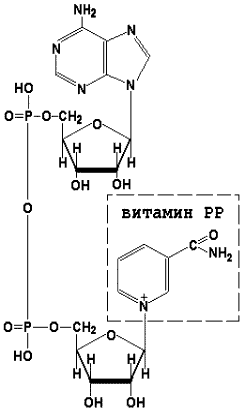
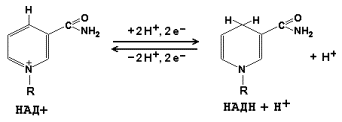
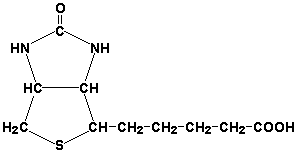
8.1.1. Как и другие функциональные белки, ферменты делятся на простые и сложные. Простые ферменты — это простые белки, они построены из аминокислот и при гидролизе распадаются только на аминокислоты. Сложные ферменты — это сложные белки, они состоят из простого белка и небелкового компонента. При их гидролизе, помимо свободных аминокислот, освобождается небелковая часть или продукты её распада. Белковая часть сложного фермента получила название апофермент, небелковая часть — кофактор. Кофакторы могут иметь разную химическую природу и отличаться по прочности связи с апоферментом. В роли кофактора могут выступать ионы различных металлов, а также другие неорганические ионы. Органические вещества неаминокислотной природы, используемые в роли кофакторов, называются коферментами. Кофермент вместе с апоферментом образуют холофермент. Кофермент + Апофермент ↔ Холофермент В некоторых случаях в условиях живой клетки равновесие в этой реакции сильно сдвинуто вправо и кофермент прочно связан со своей белковой частью, они не разделяются при выделении и очистке. Такой кофермент называется простетической группой. Следует отметить одну отличительную особенность сложных ферментов, заключающуюся в том, что ни кофактор (в том числе кофермент), ни сам по себе апофермент каталитической активностью не обладают и только их объединение в единое целое обеспечивает быстрое протекание химической реакции. Коферменты относятся к сложным органическим веществам, их молекулы значительно меньше по размеру, чем молекулы ферментов. Коферменты могут проникать через биологические мембраны, нагревание обычно не вызывает изменения их структуры. 8.1.2. Функцией кофермента является участие в катализируемой реакции, причём количество кофермента и его химическое строение внешне остаются неизменными. В действительности кофермент является одним из субстратов ферментативной реакции, т.е. выступает как косубстрат. В ходе реакции кофермент претерпевает химические превращения, в точности противоположные тем, которые происходят в субстрате. Например, в окислительно-восстановительных реакциях молекула субстрата окисляется, а молекула кофермента восстанавливается. При последующих сопряжённых реакциях изменения в коферменте протекают в обратном направлении и он воспроизводится в первоначальной форме. Таким образом, коферменты могут быть охарактеризованы как переносчики определённых атомов, электронов или химических групп на соответствующий акцептор. Строение апофермента определяет специфичность этой реакции, а строение кофермента – её тип.
|
|||||||||||||
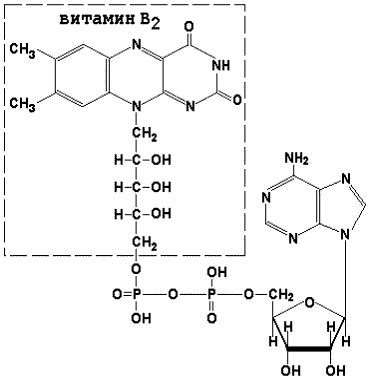
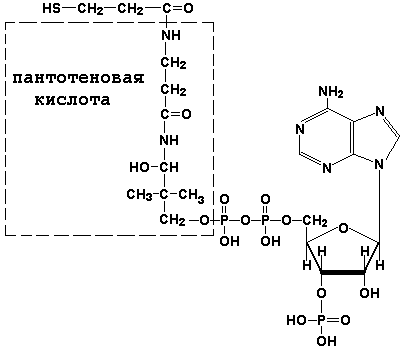
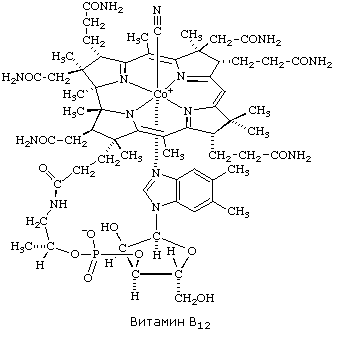
| Раздел 8.2 |
Активный центр фермента. Контактный и каталитический участки активного центра, их функции. Образование фермент-субстратного комплекса. |
|
|
|
|
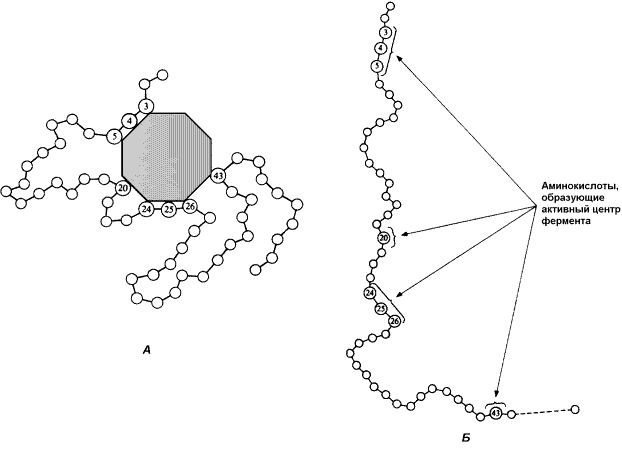
8.2.1. В процессе формирования фермент-субстратного комплекса субстрат присоединяется к специфическому участку на молекуле фермента, который называется активным центром. Активный центр – участок молекулы фермента, который связывает субстраты и от которого зависит специфичность каталитического действия ферментов; активный центр содержит функциональные группы остатков аминокислот и коферментов, пространственно сближенных и определённым образом ориентированных. Несмотря на огромное разнообразие структуры ферментов, их специфичности и механизма действия, существует ряд общих закономерностей формирования активных центров. Во-первых, на активный центр приходится относительно малая часть объёма фермента. Роль остальных аминокислотных остатков, составляющих основную массу фермента, состоит в том, чтобы обеспечить молекуле фермента правильную глобулярную форму. Во-вторых, активный центр – это сложная трёхмерная структура, и в её образовании принимают участие группы, принадлежащие разным частям линейной последовательности аминокислот. Радикалы аминокислот, образующих активный центр, оказываются вблизи друг от друга в результате формирования третичной структуры белка (рисунок 5.1). Поэтому при воздействии факторов, вызывающих денатурацию (нагревание, концентрированные кислоты и щёлочи) утрачивается конформация активного центра и фермент теряет свою активность.
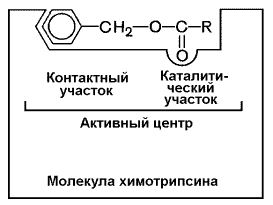
Рисунок 8.1. А. Участие аминокислотных остатков, образующих активный центр фермента, во взаимодействии с субстратом. Б. Положение этих аминокислотных остатков в первичной структуре фермента. В-третьих, активный центр имеет форму узкого углубления или щели, в которую ограничен доступ воде, за исключением тех случаев, когда вода является одним из реагирующих веществ. В этом углублении присутствует несколько полярных аминокислотных остатков, необходимых для связывания субстрата и катализа. В-четвёртых, в составе активного центра можно условно выделить две части: а) контактный или якорный участок, где происходит связывание субстрата в нужной ориентации; б) каталитический участок, обеспечивающий протекание реакции (рисунок 5.2).
Рисунок 8.2. Состав активного центра фермента (на примере химотрипсина). В-пятых, субстраты относительно слабо связываются с ферментами. В связывании и превращении субстрата принимают участие следующие группировки аминокислотных радикалов:
У сложных ферментов в формировании активных центров принимают участие также функциональные группы коферментов. В образовании фермент-субстратных комплексов принимают участие те же молекулярные взаимодействия, что и обеспечивают формирование пространственной структуры макромолекул, межклеточные контакты и другие процессы в биологических системах:
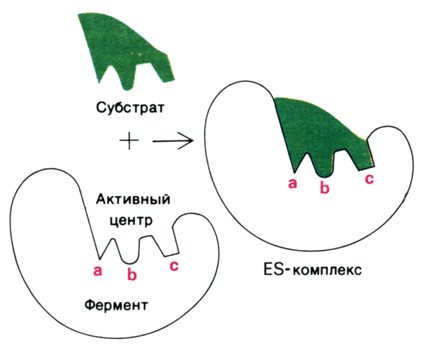
Эти три основных типа нековалентных связей различаются по своей геометрии, энергии, специфичности. 8.2.2. Cпецифичность связывания субстрата с ферментом зависит от строго определённого расположения атомов в активном центре. Субстрат входит в активный центр, если он соответствует ему по форме. Существует две модели, описывающие взаимодействие субстрата с активным центром: а) Модель жёсткого соответствия («ключ – замок»), предложена Э. Фишером в 1890 году. Активный центр считается заранее подогнанным под форму молекулы субстрата (рисунок 5.3). Эта модель не утратила своего значения для понимания некоторых свойств ферментов, например, их способности к строго определённому связыванию двух или большего числа субстратов или для объяснения кинетики насыщения субстратом.
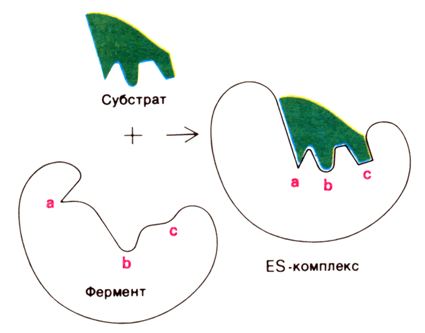
Рисунок 8.3. Взаимодействие субстрата с ферментом согласно модели жёсткого соответствия (Л.Страйер, 1984). б) Модель индуцированнного соответствия («рука – перчатка»), предложена Кошлендом в 1950-е годы. Согласно этой модели, субстрат вызывает (индуцирует) конформационные изменения фермента, и лишь в результате этих изменений аминокислотные остатки фермента принимают пространственную ориентацию, необходимую для связывания субстрата и катализа (рисунок 5.4). При этом другие аминокислотные остатки могут погрузиться вглубь молекулы фермента.
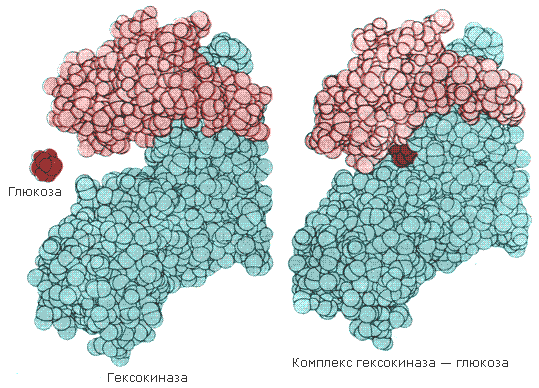
Рисунок 8.4. Взаимодействие субстрата с ферментом согласно модели индуцированного соответствия (Л.Страйер, 1984). Значение конформационных изменений, возникающих в молекуле фермента в процессе присоединения к ней субстрата можно рассмотреть на примере гексокиназы. Этот фермент катализирует фосфорилирование глюкозы в реакции с АТФ. Присоединение относительно небольшой молекулы глюкозы к активному центру гексокиназы приводит к сближению полипептидных цепей двух субъединиц, которые, как клещи, захватывают молекулу глюкозы (рисунок 8.5). По-видимому, при такой индуцированной подгонке конформации фермента к структуре субстрата молекула глюкозы также деформируется и облегчается её взаимодействие с молекулой АТФ.
Рисунок 8.5. Индуцированное соответствие при взаимодействии гекокиназы с глюкозой по данным рентгеноструктурного исследования (А.Ленинджер, 1985). |
|
| Раздел 8.3 |
Аллостерический (регуляторный) центр фермента, его функция. Роль четвертичной структуры молекулы белка в его формировании. |
|
|
|
|
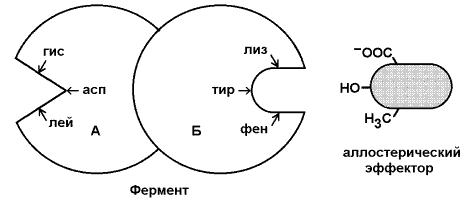
8.3.1. Многие ферменты, обладающие олигомерной структурой (хотя и не все), являются аллостерическими белками, способными изменять своё сродство к субстрату. Аллостерические ферменты, как правило, катализируют начальные реакции в многостадийных путях химических превращений в клетке. Аллостерические ферменты отличаются от остальных ферментов тем, что не подчиняются классической кинетике Михаэлиса – Ментен. Зависимость скорости реакции от концентрации субстрата у таких ферментов имеет вид S-образной кривой. Наряду с активным центром такие ферменты содержат по меньшей мере один аллостерический центр (регуляторный центр). 8.3.2. Аллостерический центр - участок молекулы фермента, способный присоединять определённые молекулы (эффекторы или модуляторы). Аллостерический центр специфичен по отношению к своему эффектору подобно тому, как активный центр специфичен по отношению к своему субстрату. Между аллостерическим центром одной из субъединиц фермента и аллостерическим эффектором могут возникать нековалентные взаимодействия (водородные, ионные и гидрофобные). Это приводит к обратимому изменению конформации остальных субъединиц молекулы фермента, в том числе изменению конформации активного центра. В результате активность фермента снижается или повышается (рисунок 8.6).
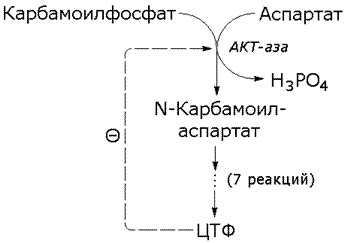
Рисунок 8.6. Схема взаимодействия аллостерического фермента и его эффектора. Аллостерический фермент состоит из двух субъединиц: А - каталитической, включающей активный центр, и Б - регуляторной, в состав которой входит аллостерический центр. Присоединение эффектора к аллостерическому центру приводит к изменению конформации активного центра. Аллостерические эффекторы бывают двух типов – активаторы и ингибиторы. Аллостерические активаторы способствуют переходу фермента из Т-конформации с низким сродством к субстрату в R-конформацию с высоким сродством к субстрату, аллостерические ингибиторы – наоборот. Если после присоединения эффектора сродство активного центра фермента к субстрату повышается, то эффектор называется аллостерическим активатором, если сродство понижается, то эффектор называется аллостерическим ингибитором. 8.3.3. Различают гомотропную и гетеротропную аллостерическую регуляцию. В случае гомотропной регуляции эффектором является субстрат. У таких ферментов аллостерический центр по своей конформации совпадает с активным, а роль аллостерического эффектора фермента выполняет молекула субстрата. Взаимодействие субстрата с активным центром одной из субъединиц аллостерического фермента повышает сродство остальных субъединиц к субстрату. Это напоминает связывание молекулы гемоглобина с кислородом. В случае гетеротропной регуляции эффектор отличается от субстрата и аллостерический центр не совпадает с активным центром. Примером может служить регуляция биосинтеза пиримидиновых нуклеотидов (рисунок 8.7).
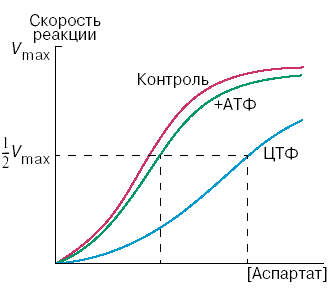
Рисунок 8.7. Схема биосинтеза пиримидиновых нуклеотидов. Начальную реакцию этого метаболического пути катализирует фермент аспартат-карбамоилтрансфераза (АКТ-аза). Конечный продукт цепи реакций – цитидинтрифосфат (ЦТФ) является аллостерическим ингибитором АКТ-азы. При увеличении концентрации ЦТФ сродство фермента к субстратам снижается, хотя максимальная скорость реакции (Vmax) остаётся неизменной (рисунок 8.8). График зависимости активности от концентрации при этом смещается вправо. В этом случае Vmax может быть достигнута при более высокой концентрации субстрата (аспартата). Ингибирующее действие ЦТФ может быть снято добавлением АТФ (субстратом промежуточных реакций биосинтеза).
Рисунок 8.8. Регуляция аспартат-карбамоилтрансферазы ЦТФ и АТФ. Таким образом, при накоплении ЦТФ в клетке скорость синтеза пиримидиновых нуклеотидов снижается и повышается при снижении концентрации ЦТФ. Так фермент обеспечивает постоянное присутствие в клетке нужных количеств цитидинтрифосфата. |
|
| Раздел 8.4 |
Понятие об активаторах и ингибиторах ферментов. Роль ионов металлов в регуляции активности ферментов. |
|
|
|
|
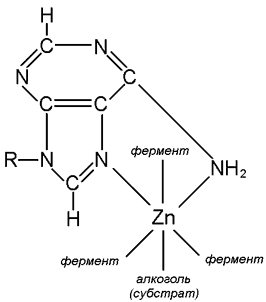
8.4.1. Как уже отмечалось, ферменты, относятся к катализаторам, активность которых может регулироваться. Поэтому через ферменты можно контролировать скорость протекающих химических реакций в организме. Регуляция активности ферментов может осуществляться путем взаимодействия с ними различных биологических компонентов или чужеродных соединений (например, лекарств и ядов), которые принято называть модификаторами или регуляторами ферментов. Под действием модификаторов на фермент реакция может ускоряться (в этом случае их называют активаторами) или замедляться (в этом случае их называют ингибиторами). 8.4.2. Активация ферментов определяется по ускорению биохимических реакций, наступающему после действия модификатора. Одну группу активаторов составляют вещества, влияющие на область активного центра фермента. К ним относятся кофакторы ферментов и субстраты. Кофакторы (ионы металлов и коферменты) являются не только обязательными структурными элементами сложных ферментов, но и по существу их активаторами. Из ионов металлов на активность многих ферментов влияют: NH4+, Na+, Mg2+, K+, Ca2+, Mn2+, Zn2+, Fe2+, Fe3+, Co2+. Ионы тяжелых металлов, как правило, оказывают ингибирующее влияние. Действие катионов в основном довольно специфично, но в большинстве случаев фермент активируется более, чем одним катионом. Наблюдается также явление антагонизма между ионами. Наиболее известен антагонизм между Na+ и К+ и между Mg2+ и Са2+. Магний является природным активатором ферментов, действующих на фосфорилированные субстраты (фосфатазы, киназы, синтетазы), но в условиях in vitro может быть заменён марганцем. Анионы в общем мало влияют на активность ферментов, и их воздействие лишено специфичности. Исключением является амилаза, активируемая хлоридами, а также, в меньшей степени, другими галогенами. Влияние активирующего иона изменяется также в зависимости от рН. Степень очистки фермента также влияет на активирующую концентрацию иона и на специфичность активации. Высокоочищенные ферменты характеризуются большей избирательностью по отношению к активирующим ионам. 8.4.3. Активирующее действие ионов металлов реализуется различными путями. Наиболее типичным механизмом является включение иона в структуру каталитического центра фермента, который без него не проявляет активности. Это типичная функция металла в роли кофермента. Другой, довольно частой функцией активирующего металла является образование связи между ферментом и субстратом, или между ферментом, коферментом и субстратом. Например, ионы Zn2+ в составе фермента алкогольдегидрогеназы образуют 2 координационные связи с молекулой кофермента НАД+, 3 координационные связи с молекулой апофермента, а шестая координационная связь присоединяет субстрат (рисунок 8.9).
Рисунок 8.9. Участие иона цинка в связывании алкогольдегидрогеназой печени кофермента и субстрата. Ионы металлов, так же как и субстраты, коферменты, их предшественники и структурные аналоги, можно использовать на практике в качестве препаратов, регулирующих активность ферментов. |
|
| Раздел 8.5 |
Виды ингибирования ферментов (обратимое и необратимое, конкурентное и неконкурентное). |
|
|
|
|
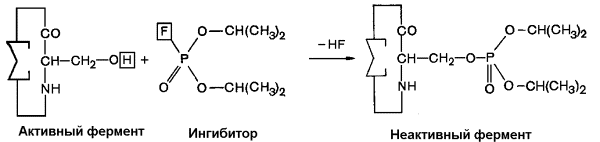
8.5.1. Ингибирование - частичное или полное торможение ферментативной реакции под действием веществ различной химической природы. Вещества, вызывающие ингибирование ферментов, называют ингибиторами. Различают обратимое и необратимое ингибирование. Если ингибитор вызывает стойкое снижение скорости реакции, то это необратимое ингибирование. При этом образуются ковалентные связи между молекулами фермента и ингибитора. Некоторые ферменты полностью ингибируются очень малыми концентрациями ионов тяжёлых металлов, например, ионов ртути (Hg2+), серебра (Ag+) и мышьяка (As+), или иодуксусной кислотой. Эти ингибиторы необратимо соединяются с SH-группами ферментов и вызывают денатурацию ферментного белка. Диизопропилфторфосфат (ДФФ) – соединение из группы нервнопаралитических отравляющих веществ. Он является ингибитором ацетилхолинэстеразы, которая инактивирует нейромедиатор ацетилхолин. ДФФ связывается с остатком аминокислоты серина в активном центре и блокирует действие фермента (рисунок 8.10). В результате ацетилхолин накапливается в синаптической щели, нервные импульсы следуют один за другим, мышца не расслабляется, и наступает паралич или смерть.
Рисунок 8.10. Необратимое ингибирование фермента ацетилхолинэстеразы диизопрропилфторфосфатом. Другим примером необратимого ингибирования может служить действие цианидов на фермент цитохромоксидазу, участвующую в окислительно-восстановительных процессах в митохондриях клеток. Отравление цианидами может привести к смерти. Если ингибитор соединяется с ферментом при помощи нековалентных связей, то возможно восстановление исходной активности фермента после удаления ингибитора, например, путём диализа. Такое ингибирование называется обратимым. 8.5.2. Обратимое ингибирование можно разделить на конкурентное и неконкурентное. Запомните особенности, характерные для конкурентного ингибирования:
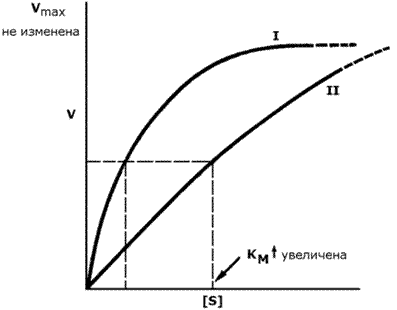
График зависимости скорости ферментативной реакции от концентрации субстрата в присутствии конкурентного ингибитора даёт такую же величину Vmax, как и в отсутствии ингибитора. Величина KM в данном случае будет увеличена, поскольку для обеспечения скорости, равной половине максимальной, в присутствии ингибитора потребуется больше субстрата. Отсюда следует, что конкурентный ингибитор препятствует образованию фермент-субстратного комплекса, но не влияет на процесс распада фермент-субстратного комплекса с образованием продуктов реакции (рисунок 8.11).
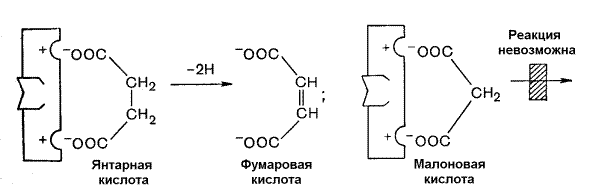
Рисунок 8.11. Влияние конкурентного ингибитора на кинетические свойства фермента. Примером конкурентного ингибирования является ингибирование фермента сукцинатдегидрогеназы малоновой кислотой (рисунок 8.12). Сукцинатдегидрогеназа катализирует реакцию дегидрирования янтарной кислоты с образованием фумаровой кислоты. Малоновая кислота, как и янтарная кислота, содержит две карбоксильные группы, но обладает более короткой углеродной цепью. Поэтому дегидрирование малоновой кислоты невозможно. Если концентрация малоновой кислоты в среде будет превышать концентрацию янтарной, то активность сукцинатдегидрогеназы снижается. Ингибирующее действие малоновой кислоты исчезает при увеличении концентрации янтарной кислоты.
Рисунок 8.12. Конкурентное ингибирование сукцинатдегидрогеназы малоновой кислотой. 8.5.3. Запомните особенности, характерные для неконкурентного ингибирования:
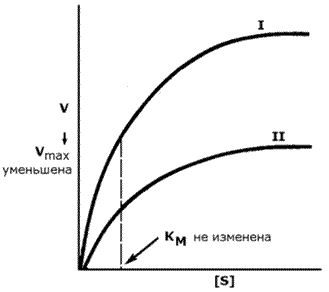
График зависимости скорости реакции от концентрации субстрата в присутствии неконкурентного ингибитора показывает сниженную величину Vmax. Субстрат не может вытеснить ингибитор из его соединения с ферментом. Величина KM в присутствии неконкурентного ингибитора не меняется. Это значит, что неконкурентный ингибитор воздействует на фермент на стадии распада фермент-субстратного комплекса, но не влияет на связывание субстрата (рисунок 8.13).
Рисунок 8.13. Влияние неконкурентного ингибитора на кинетические свойства фермента. Неконкурентные ингибиторы снижают количество молекул субстрата, которые взаимодействуют с одной молекулой фермента в единицу времени (число оборотов фермента). Ингибиторы ряда ферментов используются в медицине как химиотерапевтические препараты. Целью химиотерапии является уничтожение возбудителя болезни при помощи химических веществ, не повреждая при этом организма-хозяина. |
|
| Раздел 8.6 |
Регуляция активности ферментов путём ковалентной модификации и по принципу отрицательной обратной связи. |
|
|
|
|
8.6.1. В ряде случаев каталитическая активность ферментов может изменяться путём ковалентной модификации - в результате разрыва или образования ковалентных связей в молекуле. Существует несколько вариантов ковалентной модификации, из которых наибольший интерес представляют частичный протеолиз и регуляция путём фосфорилирования — дефосфорилирования. 8.6.2. Частичный протеолиз. Многие белки синтезируются в форме неактивных предшественников, которые затем активируются в результате специфического расщепления одной или нескольких пептидных связей. Если каталитически активный белок называется ферментом (или энзимом), то неактивный предшественник фермента называется проферментом (или зимогеном). Активация белков путем частичного протеолиза - процесс, широко распространенный в биологических системах. Вот несколько примеров.
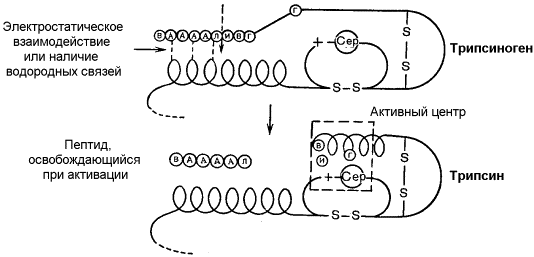
Активацию неактивных предшественников ферментов путем частичного протеолиза можно рассмотреть на примере превращения трипсиногена в трипсин. Этот процесс происходит под действием фермента энтеропептидазы в просвете двенадцатиперстной кишки и сводится к отщеплению с N-конца полипептидной цепи 6 аминокислотных остатков и соответственно укорочению полипептидной цепи (рисунок 8.14). Такое же действие на трипсиноген оказывает и активный трипсин. В результате изменения первичной структуры в молекуле профермента возникают новые нековалентные связи, изменяется конформация полипептидной цепи и формируется активный центр. В молекуле профермента активный центр отсутствует.
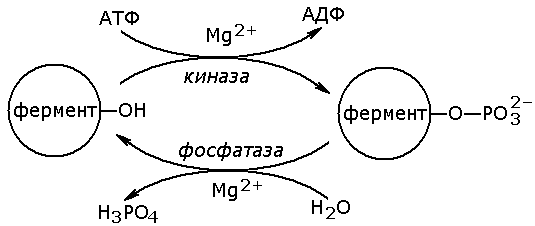
Рисунок 8.14. Схема механизма активации трипсиногена быка. Физиологический смысл выработки пищеварительных ферментов в форме проферментов заключается в том, что в противном случае ферменты могли бы оказывать свой эффект на клеточные белки слизистой желудка и поджелудочной железы, вызывая разрушение этих клеток. Такое разрушение клеток может наблюдаться, например, при панкреатите, когда активация трипсина происходит непосредственно в поджелудочной железе. Фосфорилирование – дефосфорилирование ферментов – присоединение или отщепление фосфатной группы. В отличие от частичного протеолиза, это обратимое изменение каталитической активности ферментов. Такие ферменты могут существовать в двух формах – фосфорилированной и дефосфорилированной. В зависимости от конкретного случая, одна из этих форм будет обладать более высокой, а другая – более низкой каталитической активностью. Фосфорилированию обычно подвергаются остатки серина, реже тирозина или треонина. Донором фосфатной группы является молекула АТФ. Фосфорилирование происходит избирательно и затрагивает лишь небольшое число аминокислотных остатков, не обязательно в активном центре фермента. Присоединение фосфата приводит к изменению конформации фермента и его активности. Фосфатные группы, связанные с остатками аминокислот, удаляются путём гидролиза с образованием неорганической фосфорной кислоты. Фосфорилирование и дефосфорилирование катализируется протеинкиназами и протеинфосфатазами соответственно (рисунок 8.15). Активность протеинкиназ и протеинфосфатаз находится под гормональным контролем и регулируется также нервной системой.
Рисунок 8.15. Ковалентная модификация, осуществляемая путем фосфорилирования — дефосфорилирования. Примером фермента, активность которого регулируется путём обратимого фосфорилирования, является гликогенфосфорилаза, участвующая в распаде гликогена в клетках печени и мышц. Неактивная форма фермента (дефосфорилированная) превращается в активную форму (фосфорилированную) при помощи другого фермента – киназы фосфорилазы. Реакцию дефосфорилирования катализирует фосфатаза фосфорилазы которая инактивирует фосфорилазу. 8.6.3. Регуляция по принципу отрицательной обратной связи. В результате аллостерических механизмов и ковалентной модификации происходит изменение активности уже имеющихся в клетке молекул фермента. Существуют также механизмы, влияющие на скорость реакций обмена веществ путём изменения количества молекул ферментативного белка в клетке. В настоящее время установлено, что синтез и распад ферментов, как и других белков, происходит в организме непрерывно. У взрослого здорового человека в условиях динамического равновесия процессы синтеза и распада имеют одинаковую скорость, благодаря чему общее содержание фермента не изменяется во времени. Для каждого фермента характерна своя скорость распада. В большинстве случаев полное прекращение синтеза фермента привело бы к исчезновению 50% молекул фермента за несколько дней, но некоторые ферменты обновляются значительно быстрее. Скорость синтеза фермента может варьировать от нуля до максимума, тогда как скорость распада представляется постоянной. Таким образом, любое вещество, влияющее на скорость синтеза фермента, способно оказать существенное воздействие на регуляцию обмена веществ путем изменения соотношения ферментов в организме. В основе многих гормональных воздействий на обмен веществ у человека лежат, как было установлено, именно такие контролирующие влияния на выработку каталитически активных белков.
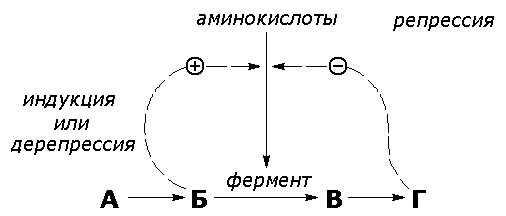
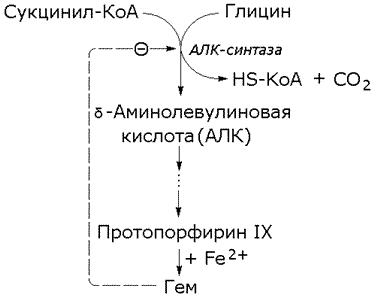
Рисунок 8.16. Регуляция синтеза фермента. Вещество, которое избирательно препятствует синтезу определенного фермента, называется репрессором. При помощи механизма репрессии конечные продукты реакций обмена веществ могут регулировать процесс их собственного образования по принципу обратной связи. Было доказано, что в некоторых системах накопление метаболитов, образующихся в итоге цепи последовательных реакций, предотвращает синтез одного из ферментов, функционирующего в начале этой цепи (рисунок 8.16). Продукт реакции в таком случае действует как специфический репрессор синтеза этого фермента предотвращая как ненужное потребление субстратов, вовлекаемых в реакции данной метаболической цепи, так и бесполезный расход энергии и аминокислот, необходимых для образования каталитически активного белка. Примером того, как конечные продукты цепи химических реакций способны замедлять синтез ферментных белков, катализирующих начальные стадии процесса (то есть снижать количество молекул этих ферментов), может служить регуляция синтеза гемоглобина в клетках кроветворных органов. По мере накопления гема в этих клетках подавляется синтез фермента, катализирующего первую реакцию синтеза гема (рисунок 8.17). Тем самым предупреждается избыточное накопление гемоглобина в клетке.
Рисунок 8.17. Регуляция синтеза гема по механизму репрессии на уровне фермента, катализирующего начальную реакцию этого метаболического пути. Явление, противоположное репрессии, известно под названиями индукция фермента или дерепрессия. В типичном случае субстрат определенного фермента способен индуцировать синтез этого фермента, что в свою очередь стимулирует потребление данного субстрата. Воздействуя на механизм синтеза фермента, индуктор, вероятно, прямо или косвенно противодействует репрессору. Соотношение между репрессором (конечным продуктом) и индуктором (субстратом) определяет, таким образом, количество ключевых ферментов и обеспечивает приспособление последовательности метаболических реакций к количеству метаболитов, поступающих в клетки организма с пищей. Как и в случае регуляторных ферментов, лишь немногие ключевые ферменты способны реагировать подобным образом на изменение физиологических потребностей. Такие ферменты называютиндуцибельными (или адаптивными); ферменты, содержание которых в таких условиях не изменяется, называют конститутивными; они составляют постоянное содержимое клетки. У человека на адаптивные ферменты, вероятно, в большей мере влияют эндокринные факторы, нежели промежуточные продукты реакций обмена веществ. Так, гормоны коры надпочечниковглюкокортикоиды стимулируют синтез ферментов, участвующих в образовании сахара крови (глюкозы), тогда как гормон поджелудочной железы инсулин противодействует этому. Глюкокортикоиды прямо или косвенно играют роль индукторов ферментов, когда как инсулин усиливает процесс репрессии. От определяемой противоположными воздействиями индукции и репрессии уровня синтеза ферментов зависит физиологическая регуляция содержания глюкозы в крови этими противоборствующими эндокринными системами. |
|
| Раздел 8.7 |
Основные компартменты клетки. Локализация ферментов в клетке и тканях. Понятие об энзимопатиях. |
|
|
|
|
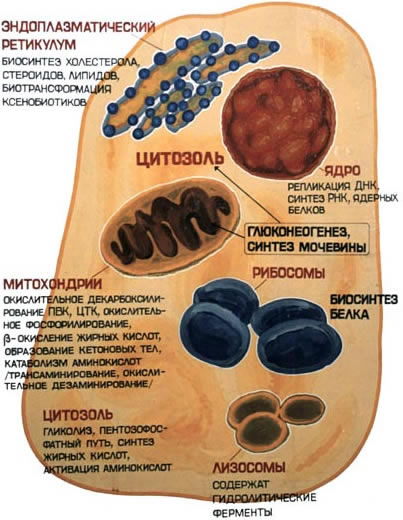
8.7.1. В клеточном содержимом ферменты распределены не хаотически, а строго упорядоченно. При помощи внутриклеточных мембран клетка разделена на отсеки или компартменты(рисунок 8.18). В каждом из них осуществляются строго определенные биохимические процессы и сосредоточены соответствующие ферменты или полиферментные комплексы. Вот несколько характерных примеров.
Рисунок 8.18. Внутриклеточное распределение ферментов различных метаболических путей. В лизосомах сосредоточены преимущественно разнообразные гидролитические ферменты. Здесь протекают процессы расщепления сложных органических соединений на их структурные компоненты. В митохондриях находятся сложные системы окислительно-восстановительных ферментов. Ферменты активирования аминокислот распределены в гиалоплазме, но они же есть и в ядре. В гиалоплазме присутствуют многочисленные метаболоны гликолиза, структурно объединенные с таковыми пентозофосфатного цикла, что обеспечивает взаимосвязь дихотомического и апотомического путей распада углеводов. В то же время ферменты, ускоряющие перенос аминокислотных остатков на растущий конец полипептидной цепи и катализирующие некоторые другие реакции в процессе биосинтеза белка, сосредоточены в рибосомальном аппарате клетки. В клеточном ядре локализованы в основном нуклеотидилтрансферазы, ускоряющие реакцию переноса нуклеотидных остатков при новообразовании нуклеиновых кислот. 8.7.2. Распределение ферментов по субклеточным органеллам изучают после предварительного фракционирования клеточных гомогенатов путем высокоскоростного центрифугирования, определяя содержание ферментов в каждой фракции. Локализацию данного фермента в ткани или клетке часто удается установить in situ гистохимическими методами («гистоэнзимология»). Для этого тонкие (от 2 до 10 мкм) срезы замороженной ткани обрабатывают раствором субстрата, к которому специфичен данный фермент. В тех местах, где находится фермент, образуется продукт катализируемой этим ферментом реакции. Если продукт окрашен и нерастворим, он остается на месте образования и позволяет локализовать фермент. Гистоэнзимология дает наглядную и в известной мере физиологичную картину распределения ферментов. Ферментные системы ферментов, сосредоточенные во внутриклеточных структурах, тонко координированы друг с другом. Взаимосвязь катализируемых ими реакций обеспечивает жизнедеятельность клеток, органов, тканей и организма в целом. При исследовании активности различных ферментов в тканях здорового организма можно получить картину их распространения. Оказывается, что некоторые ферменты широко распространены во многих тканях, но в разных концентрациях, а другие очень активны в экстрактах, полученных из одной или нескольких тканей, и практически отсутствуют в остальных тканях организма.
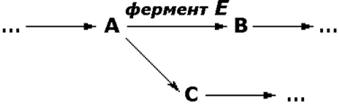
Рисунок 8.19. Относительная активность некоторых ферментов в тканях человека, выраженная в процентах от активности в ткани с максимальной концентрацией данного фермента (Мосс, Баттерворт, 1978). 8.7.3. Понятие об энзимопатиях. В 1908 году английский врач Арчибальд Гаррод высказал предположение, что причиной ряда заболеваний может являться отсутствие какого-либо из ключевых ферментов, участвующих в обмене веществ. Он ввёл понятие "inborn errors of metabolism" (врождённый дефект обмена веществ). В дальнейшем эта теория была подтверждена новыми данными, полученными в области молекулярной биологии и патологической биохимии. Информация о последовательности аминокислот в полипептидной цепи белка записана в соответствующем участке молекулы ДНК в виде последовательности тринуклеотидных фрагментов - триплетов или кодонов. Каждый триплет кодирует определённую аминокислоту. Такое соответствие называется генетическим кодом. Причём некоторые аминокислоты могут быть закодированы при помощи нескольких кодонов. Существуют также специальные кодоны, являющиеся сигналами для начала синтеза полипептидной цепи и его прекращения. К настоящему времени генетический код полностью расшифрован. Он является универсальным для всех видов живых организмов. Реализация информации, заложенной в молекуле ДНК, включает несколько этапов. Сначала в клеточном ядре в процессе транскрипции синтезируется матричная РНК (мРНК), поступающая в цитоплазму. В свою очередь, мРНК служит матрицей для трансляции - синтеза полипептидных цепей на рибосомах. Таким образом, природа молекулярных болезней определяется нарушением структуры и функции нуклеиновых кислот и контролируемых ими белков. 8.7.4. Поскольку информация о структуре всех белков клетки содержится в последовательности нуклеотидов ДНК, а каждая аминокислота определяется триплетом нуклеотидов, изменение первичной структуры ДНК может в конечном счёте оказать глубокое влияние на синтезируемый белок. Подобные изменения происходят за счёт ошибок репликации ДНК, когда одно азотистое основание заменяется другим, либо в результате действия радиации или при химической модификации. Все возникшие таким образом наследуемые дефекты называются мутациями. Они могут приводить к неправильному считыванию кода и делеции (выпадению) ключевой аминокислоты, замене одной аминокислоты другой, преждевременной остановке белкового синтеза или добавлению аминокислотных последовательностей. Учитывая зависимость пространственной упаковки белка от линейной последовательности в нём аминокислот, можно полагать, что подобные дефекты способны изменить структуру белка, а значит, и его функцию. Тем не менее, многие мутации обнаруживаются только в лабораторных условиях и не оказывают вредного воздействия на функции белка. Таким образом, ключевым моментом является локализация изменений в первичной структуре. Если положение замененной аминокислоты окажется критическим для формирования третичной структуры и образования каталитического центра фермента, то мутация является серьёзной и может проявиться как заболевание. Последствия недостаточности одного фермента в цепи реакций обмена веществ могут проявляться по-разному. Предположим, что превращение соединения A в соединение B катализирует фермент Е и что соединение C встречается на альтернативном пути превращений (рисунок 8.20):
Рисунок 8.20. Схема альтернативных путей биохимических превращений. Последствиями недостаточности фермента могут быть следующие явления:
Если метаболическое превращение в целом регулируется по принципу обратной связи конечным продуктом, то эффекты двух последних типов аномалий будут более значительными. Так, например, при порфириях (врождённых нарушениях синтеза гема) устраняется подавляющего эффекта гема на начальные реакции синтеза, что приводит к образованию избыточных количеств промежуточных продуктов метаболического пути, которые обладают токсическим действием на клетки кожи и нервной системы. Факторы внешней среды могут усиливать или даже полностью определять клинические проявления некоторых врождённых нарушений обмена веществ. Например, у многих пациентов с недостаточностью глюкозо-6-фосфатдегидрогеназы заболевание начинается только после приёма таких лекарственных средств, как примахин. В отсутствие контактов с лекарственными средствами такие люди производят впечатление здоровых. 8.7.5. О недостаточности фермента обычно судят косвенно по повышению концентрации исходного вещества, которое в норме подвергается превращениям под действием данного фермента (например, фенилаланин при фенилкетонурии). Прямое определение активности таких ферментов проводят только в специализированных центрах, но по возможности диагноз следует подтверждать этим методом. Пренатальная (дородовая) диагностика некоторых врождённых нарушений метаболизма возможна путём иследования клеток амниотической жидкости, полученных на ранних стадиях беременности и культивируемых in vitro. Некоторые врождённые нарушения метаболизма поддаются лечению путём доставки в организм недостающего метаболита или путём ограничения поступления в желудочно-кишечный тракт предшественников нарушенных процессов обмена веществ. Иногда могут быть удалены накапливающиеся продукты (например, железо при гемохроматозе). |
|
| Раздел 8.8 |
Структурно-функциональная организация ферментов: изоферменты и мультиферменты: их биологическая роль. |
||||||||||||
|
|
|||||||||||||
|
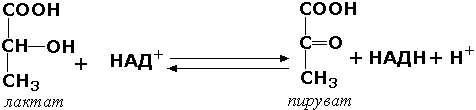
8.8.1. Изоферментами или изозимами называют множественные формы ферментов, которые существуют у одного и того же вида, в одной и той же ткани, и даже в одной и той же клетке. Все эти формы фермента катализируют одну и ту же реакцию, но различаются по своим кинетическим свойствам, а также по первичной структуре. Изоферменты играют регуляторную роль в обмене веществ и позволяют метаболизму в разных тканях лучше приспосабливаться к действию внутренних и внешних факторов. Примером фермента, у которого были обнаружены такие формы, может служить лактатдегидрогеназа (L-лактат:НАД+-оксидоредуктаза, КФ 1.1.1.27), катализирующая обратимую окислительно-восстановительную реакцию:
Лактатдегидрогеназа (ЛДГ) присутствует в тканях животных в виде пяти разных изоферментов, которые различаются на уровне четвертичной структуры. Молекула ЛДГ состоит из четырех протомеров двух типов, Н (от англ. heart - сердце) и М (от англ. muscle - мышца), которые различаются по аминокислотному составу и последовательности аминокислот. Каталитической активностью обладает только тетрамерная молекула. Протомеры могут быть скомпонованы следующими способами:
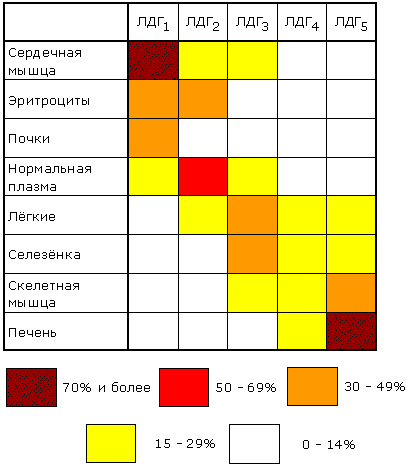
Изоферменты сывороточной лактатдегидрогеназы могут быть обнаружены с помощью электрофореза при рН 8,6. При данном значении рН изозимы несут разный заряд и распределяются на электрофореграмме в пяти разных местах. Наибольшим отрицательным зарядом обладает изозим ЛДГ1. Распределение изоферментов ЛДГ (изоферментный спектр) в тканях также отличается. Так, изоформа ЛДГ, содержащая четыре М-субъединицы, преобладает в печени и скелетной мышце, а изоформа, состоящая из четырех Н-субъединиц, преобладает в миокарде (рисунок 8.21).
Рисунок 8.21. Относительное содержание изоферментов ЛДГ (в процентах от суммарной активности) в некоторых тканях человека (Мосс, Баттерворт, 1978). 8.8.2. Мультиферменты (мультэнзимы) - надмолекулярные комплексы, в состав которых входят ферменты, катализирующие последовательные стадии превращения субстрата. Например, для в реакциях превращения метаболита A в метаболит D :
комплекс ферментов Е1, Е2, Е3 является мультиферментом. Объединение нескольких ферментов в один комплекс имеет важное преимущество: резко сокращаются расстояния, на которые молекулы промежуточных продуктов должны перемещаться от фермента к ферменту. Поэтому суммарная скорость таких метаболических путей довольно высока. Примером мультэнзима может служить пируватдегидрогеназный комплекс, находящийся в митохондриях и катализирует последовательные реакции окислительного декарбоксилирования пирувата:
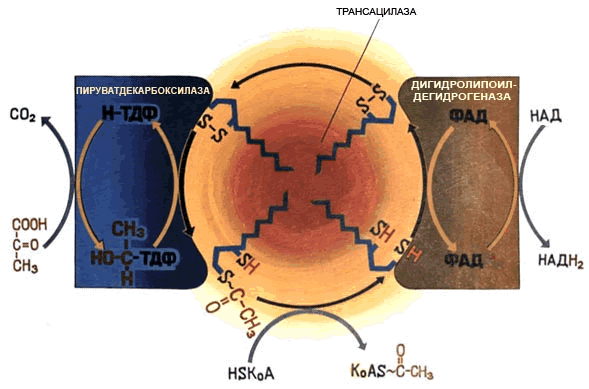
Пируватдегидрогеназный комплекс состоит из трёх ферментов: пируватдекарбоксилазы, трансацилазы и дигидролипоилдегидрогеназы (рисунок 8.22).
Рисунок 8.22. Схема организации и работы пируватдегидрогеназного комплекса. В промежуточных реакциях участвует пять коферментов:
Регуляторным ферментом комплекса является пируватдекарбоксилаза, активность которой (и всего комплекса в целом) снижается при высокой концентрации АТФ в клетке. |
|||||||||||||
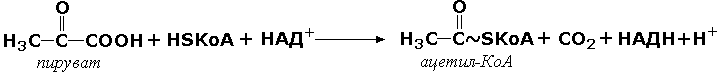
| Раздел 8.8 |
Определение ферментов в плазме крови с целью диагностики заболеваний. Ферменты как аналитические реагенты. |
||||||||||||||||||||||
|
|
|||||||||||||||||||||||
|
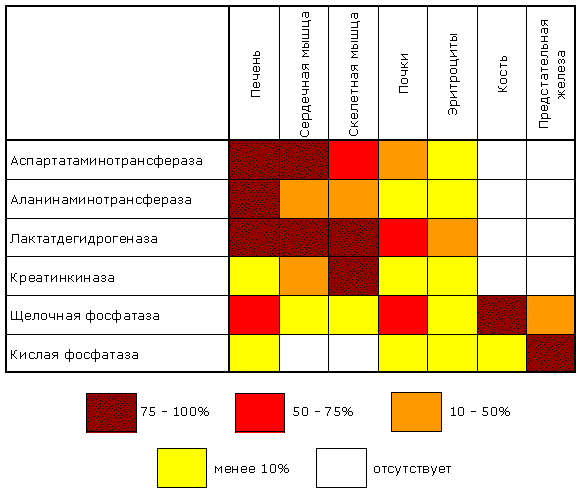
8.9.1. Некоторые ферменты, проферменты и их субстраты в норме постоянно циркулируют в крови человека и выполняют физиологические функции. Примерами таких ферментов плазмы являются липопротеинлипаза, псевдохолинэстераза, а также проферменты компонентов систем свертывания крови и растворения кровяного сгустка. Эти ферменты называютсясекреторными, большинство из них синтезируется в печени и секретируются в кровь. Концентрация таких энзимов в крови либо такая же, как в тканях, либо более высокая. Кроме секреторных ферментов, в плазме крови присутствуют ферменты, которые не выполняют в крови никаких известных физиологических функций. Их субстраты в плазме обычно не обнаруживаются, и в норме их концентрация в крови человека почти в миллион раз ниже, чем в тканях. Такие ферменты называются индикаторными. Появление их в плазме крови в повышенных концентрациях указывает на повышенную скорость разрушения тканей. Таким образом, измерение в крови уровня индикаторных ферментов плазмы дает врачу ценную диагностическую и прогностическую информацию. Индикаторные ферменты обычно обнаруживаются в плазме в малых количествах, и появляются в ней вследствие постоянно протекающих процессов разрушения клеток организма. Появление этих ферментов в плазме в повышенных концентрациях указывает на повышенную скорость разрушения тканей. Поступлением в плазму значительных количеств мышечных ферментов сопровождается и выполнение тяжелой физической работы. Для клеток разных органов характерен свой набор ферментов, поэтому повышение в крови активности того или иного фермента может указывать на заболевание соответствующего органа. В клинической практике используется количественное определение различных индикаторных ферментов плазмы. В клетках поджелудочной железы вырабатываются ферменты трипсин (трипсиноген), липаза и амилаза; при остром воспалении поджелудочной железы (острый панкреатит) активность этих ферментов в крови возрастает; повышенная активность амилазы обнаруживается и в моче. Довольно часто в целях диагностики проводят анализ изоферментного спектра некоторых ферментов, в частности ЛДГ. В клетках миокарда преобладает изофермент ЛДГ1. При заболеваниях, связанных с повреждением сердечной мышцы, концентрация и активность этого изофермента в плазме крови значительно возрастает. При некоторых заболеваниях печени (в том числе инфекционной природы) в крови возрастает содержание ЛДГ4 и ЛДГ5 - изоферментов, характерных для клеток печени. В настоящее время для получения этой ценной диагностической и прогностической информации в большинстве случаев используются автоанализаторы. В таблице 8.1 приведен перечень ферментов, активность которых чаще всего исследуют в диагностической энзимологии. Таблица 6.1. Основные ферменты сыворотки, используемые в клинической диагностике.
8.9.2. Высокая специфичность ферментов позволяет использовать их для обнаружения и количественного определения нормальных и патологических компонентов крови и мочи. Например, с помощью фермента уреазы, действующего только на мочевину, можно проводить определение этого вещества в биологических жидкостях. Фермент глюкозооксидаза применяется для определения глюкозы в крови и моче у больных сахарным диабетом. |
|||||||||||||||||||||||
ОБНОВЛЕНИЯ
08 июня 2019, 16:18:59
08 июня 2019, 16:10:24
07 июня 2019, 22:59:05
09 мая 2019, 16:13:23
27 апреля 2019, 16:00:00
24 апреля 2019, 01:02:18
17 апреля 2019, 22:26:23
17 апреля 2019, 22:21:37
17 апреля 2019, 22:07:42
17 апреля 2019, 22:06:28
ПОДПИСАТЬСЯ НА РАССЫЛКУ
ПРЕДМЕТЫ
- Анатомия
- Акушерство и гинекология
- БЖД, медицина катастроф
- Биохимия
- Биология
- Гистология
- Гигиена
- Генетика
- Диетология
- Дерматовенерология
- Инфекционные болезни
- Культурология
- Лабораторная диагностика
- Летняя практика
- Лучевая диагностика
- Медицинская информатика
- Микробиология
- Неврология
- Общественное здоровье
- Общий уход
О НАС
«Dendrit» - информационный портал для медицинских работников, студентов медицинских ВУЗов, исследователей и пациентов.
Ваш источник новостей и знаний о здоровье.
dendrit.ru © 2014-2024
