Dendrit
Новости, статьи, учебные материалы по медицине
ПОДПИСАТЬСЯ НА РАССЫЛКУ
Структура и биологическая роль нуклеотидов, нуклеиновых кислот. Репликация ДНК и транскрипция.
Тема: «СТРУКТУРА И БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕОТИДОВ И НУКЛЕИНОВЫХ КИСЛОТ. МАТРИЧНЫЕ БИОСИНТЕЗЫ (РЕПЛИКАЦИЯ ДНК И ТРАНСКРИПЦИЯ)»
1. Нуклеопротеины: разновидности, особенности аминокислотного состава апопротеинов. Формулы пуриновых и пиримидиновых азотистых оснований, нуклеозидов, нуклеотидов.
2. Понятие о первичной и вторичной структуре нуклеиновых кислот. Типы химических связей, стабилизирующих структуру нуклеиновых кислот.
3. Особенности нуклеотидного состава ДНК и РНК. Правила Чаргаффа. Сравнительная характеристика ДНК и РНК (молекулярная масса, пространственная структура, локализация в клетке, биологическая роль).
4. Понятие о матричных биосинтезах. Основные типы матричных биосинтезов.
5. Основные принципы и особенности процесса репликации ДНК (матрица, важнейшие ферменты, субстраты реакций, источники энергии, направление полимеризации полинуклеотидных цепей).
6. Нарушение комплементарности цепей ДНК в результате ошибок репликации. Повреждения ДНК под действием физических и химических факторов. Механизмы репарации ДНК.
7. Основные принципы и особенности процесса транскрипции (матрица, важнейшие ферменты, субстраты реакций, источники энергии, направление полимеризации полинуклеотидных цепей).
8. Ковалентная модификация (процессинг) матричной РНК: основные операции.
| Раздел 4.1 | Нуклеиновые кислоты, их состав. Строение азотистых оснований, нуклеозидов и нуклеотидов. |
|
|
|
|
4.1.1. Нуклеиновыми кислотами или полинуклеотидами называются высокомолекулярные вещества, состоящие из нуклеотидов, соединённых в цепь 3', 5'-фосфодиэфирными связями. Каждый нуклеотид состоит из азотистого основания, углевода (пентозы) и остатка фосфорной кислоты.
Азотистые основания, входящие в состав нуклеотидов, имеют следующее строение:
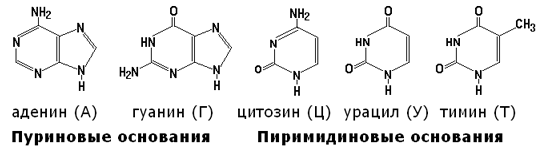 Углеводы представлены рибозой и дезоксирибозой:
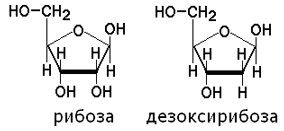 4.1.2. Азотистое основание и пентоза, соединённые N-гликозидной связью, образуют нуклеозид. Если в качестве пентозы в нуклеозиде присутствует рибоза, то это рибонуклеозид, а если дезоксирибоза - то это дезоксирибонуклеозид.
|
|
4.1.3. Нуклеотиды представляют собой фосфорилированные нуклеозиды. Остаток фосфорной кислоты, как правило, присоединяется к гидроксильной группе пентозы в 5'-положении при помощи сложноэфирной связи. Примеры:
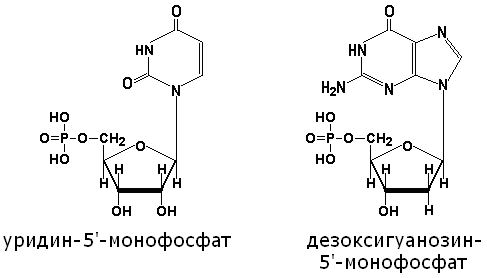
В клетках встречаются также нуклеозиддифосфаты и нуклеозидтрифосфаты, содержащие соответственно два и три остатка фосфорной кислоты. Биологическая роль этих соединений будет рассматриваться в дальнейшем.
Выучите формулы нуклеотидов и научитесь давать им правильные названия (таблица 3.3.).
Кроме перечисленных, известны минорные нуклеотиды (редко встречающиеся) . Они содержат, как правило, метилированные производные вышеприведённых главных азотистых оснований. Минорные основания присутствуют в составе некоторых разновидностей рибонуклеиновых кислот. Роль этих оснований заключается, очевидно, в защите молекулы нуклеиновой кислоты от действия гидролитических ферментов.
| Азотистое основание | Рибонуклеозиды и их производные | Дезоксирибонуклеозиды и их производные | ||||||
|---|---|---|---|---|---|---|---|---|
| нуклеозид (азот.осн. + рибоза) | нуклеотид (нуклеозид + фосфат) | нуклеозид-дифосфат | нуклеозид-трифосфат | нуклеозид (азот.осн. + рибоза) | нуклеотид (нуклеозид + фосфат) | нуклеозид-дифосфат | нуклеозид-трифосфат | |
| аденин | аденозин | аденозин-5'-монофосфат (АМФ, адениловая кислота) | аденозин-5'-дифосфат (АДФ) | аденозин-5'-трифосфат (АТФ) | дезоксиаденозин | дезоксиаденозин-5'-монофосфат (дАМФ, дезоксиадениловая кислота) | дезоксиаденозин-5'-дифосфат (дАДФ) | дезоксиаденозин-5'-трифосфат (дАТФ) |
| гуанин | гуанозин | гуанозин-5'-монофосфат(ГМФ, гуаниловая кислота) | гуанозин-5'-дифосфат (ГДФ) | гуанозин-5'-трифосфат (ГТФ) | дезоксигуанозин | дезоксигуанозин-5'-монофосфат (дГМФ, дезоксигуаниловая кислота) | дезоксигуанозин-5'-дифосфат (дГДФ) | дезокси-гуанозин-5'-трифосфат (дГТФ) |
| цитозин | цитидин | цитидин-5'-монофосфат (ЦМФ, цитидиловая кислота) | цитидин-5'-дифосфат (ЦДФ) | цитидин-5'-трифосфат(ЦТФ) | дезоксицитидин | дезоксицитидин-5'-монофосфат(дЦМФ, дезоксицитидиловая кислота) | дезоксицитидин-5'-дифосфат (дЦДФ) | дезоксицитидин-5'-трифосфат (дЦТФ) |
| урацил | уридин | уридин-5'-монофосфат (УМФ, уридиловая кислота) | уридин-5'-дифосфат (УДФ) | уридин-5'-трифосфат (УТФ) | - | - | - | - |
| тимин* | - | - | - | - | (дезокси)-тимидин* | (дезокси)тимидин-5'-монофосфат (дТМФ, дезокситимидиловая кислота) | (дезокси)тимидин-5'-дифосфат (дТДФ) | (дезокси)тимидин-5'-трифосфат (дТТФ) |
* В названиях производных тимина приставка дезокси-, как правило, опускается (т.к. это основание содержится исключительно в дезоксирибонуклеотидах)
| Раздел 4.2 |
Первичная структура нуклеиновых кислот. Особенности строения ДНК и РНК и их функции в клетке. |
|
|
|
|
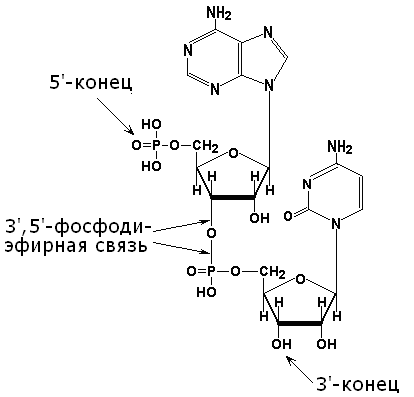
4.2.1. Первичной структурой нуклеиновых кислот называется последовательность расположения мононуклеотидов в цепи ДНК или РНК. Первичная структура нуклеиновых кислот стабилизируется 3',5'-фосфодиэфирными связями. Эти связи образуются при взаимодействии гидроксильной группы в 3'-положении пентозного остатка каждого нуклеотида с фосфатной группой соседнего нуклеотида (рисунок 3.2), Таким образом, на одном конце полинуклеотидной цепи имеется свободная 5'-фосфатная группа (5'-конец), а на другом - свободная гидроксильная группа в 3'-положении (3'-конец). Нуклеотидные последовательности принято записывать в направлении от 5'-конца к 3'-концу.
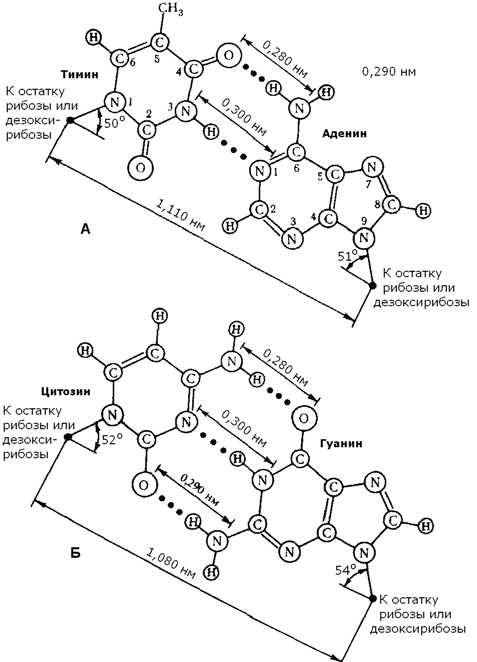
Рисунок 4.2. Структура динуклеотида, в состав которого входят аденозин-5'-монофосфат и цитидин-5'-монофосфат. 4.2.2. ДНК (дезоксирибонуклеиновая кислота) содержится в клеточном ядре и имеет молекулярную массу порядка 1011 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, тимин, углевод дезоксирибоза и остатки фосфорной кислоты. Содержание азотистых оснований в молекуле ДНК определяют правила Чаргаффа: 1) количество пуриновых оснований равно количеству пиримидиновых (А + Г = Ц + Т) ; 2) количество аденина и цитозина равно количеству тимина и гуанина соответственно (А = Т; Ц = Г) ; 3) ДНК, выделенные из клеток различных биологических видов, отличаются друг от друга величиной коэффициента специфичности: (Г + Ц) /(А + Т) Эти закономерности в строении ДНК объясняются следующими особенностями её вторичной структуры: 1) молекула ДНК построена из двух полинуклеотидных цепей, связанных между собой водородными связями и ориентированных антипараллельно (то есть 3'-конец одной цепи расположен напротив 5'-конца другой цепи и наоборот); 2) водородные связи образуются между комплементарными парами азотистых оснований. Аденину комплементарен тимин; эта пара стабилизируется двумя водородными связями. Гуанину комплементарен цитозин; эта пара стабилизируется тремя водородными связями (см. рисунок б) . Чем больше в молекуле ДНК пар Г-Ц, тем больше её устойчивость к действию высоких температур и ионизирующего излучения;
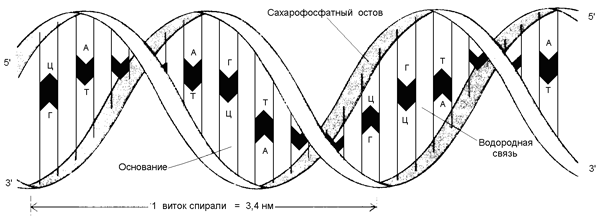
Рисунок 3.3. Водородные связи между комплементарными азотистыми основаниями. 3) обе цепи ДНК закручены в спираль, имеющую общую ось. Азотистые основания обращены внутрь спирали; кроме водородных, между ними возникают также гидрофобные взаимодействия. Рибозофосфатные части расположены по периферии, образуя остов спирали (см. рисунок 3.4).
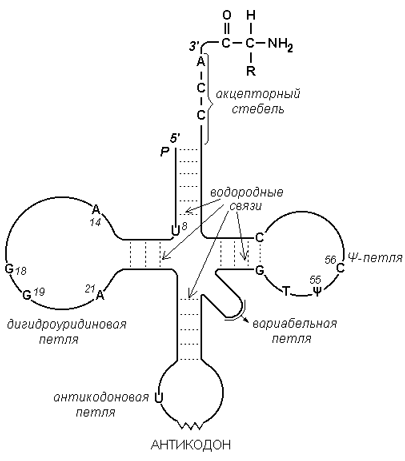
Рисунок 3.4. Схема строения ДНК. 4.2.3. РНК (рибонуклеиновая кислота) содержится преимущественно в цитоплазме клетки и имеет молекулярную массу в пределах 104 - 106 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, урацил, углевод рибоза и остатки фосфорной кислоты. В отличие от ДНК, молекулы РНК построены из одной полинуклеотидной цепи, в которой могут находиться комплементарные друг другу участки (рисунок 3.5). Эти участки могут взаимодействовать между собой, образуя двойные спирали, чередующиеся с неспирализованными участками.
Рисунок 3.5. Схема строения транспортной РНК. По особенностям структуры и функции различают три основных типа РНК: 1) матричные (информационные) РНК (мРНК) передают информацию о структуре белка из клеточного ядра на рибосомы; 2) транспортные РНК (тРНК) осуществляют транспорт аминокислот к месту синтеза белка; 3) рибосомальные РНК (рРНК) входят в состав рибосом, участвуют в синтезе белка. |
|
| Раздел 4.3 | Общая характеристика |
|
|
|
|
4.3.1. Матричный биосинтез – процесс сборки новых макромолекул из мономеров, последовательность которых запрограммирована с помощью нуклеиновых кислот. Молекулы, используемые в качестве программы в матричном биосинтезе, называют матрицами.
Тремя главными матричными биосинтезами, присущими всем без исключения живым организмам, являются репликация ДНК, транскрипция и трансляция.
Связь этих процессов отражена в основном постулате молекулярной биологии: направление потока информации от генотипа к фенотипу: ДНК → РНК → белок (стрелки обозначают направление передачи информации).
4.3.2. Кроме того, для некоторых видов вирусов характерны ещё два вида матричных синтезов:
4.3.3. Попытаемся сформулировать общие закономерности, характерные для всех матричных биосинтезов.
|
|
| Раздел 4.4 |
Репликация и репарация ДНК. |
|
|
|
|
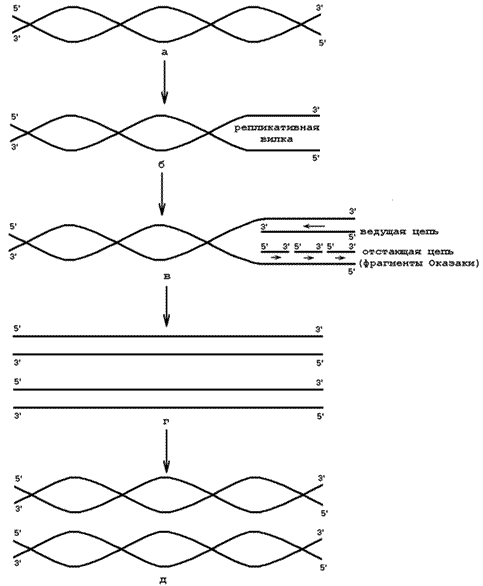
4.4.1. Репликация – процесс самоудвоения ДНК, или биосинтез дочерней молекулы ДНК, полностью идентичной исходной молекуле (матрице). Локализация процесса – клеточное ядро. Основные принципы репликации ДНК:
4.4.2. Условия, необходимые для репликации ДНК:
Рисунок 8.1. Репликация ДНК (объяснения в тексте). 1) Матрица – молекула ДНК (рисунок 26.1, а); 2) Расплетающие белки – разрывают водородные связи между комплементарными основаниями двойной спирали ДНК, в результате чего образуется репликативная вилка (рисунок 26.1, б); 3) ДНК-связываюшие белки - присоединяются к разделившимся цепям ДНК и препятствуют их обратному воссоединению; 6) ДНК-полимераза – фермент, который формирует из нуклеозидтрифосфатов новые полинуклеотидные цепи за счёт образования 3’,5’-фосфодиэфирных связей. Источником энергии служат макроэргические связи нуклеозидтрифосфатов. На одной ветви репликативной вилки синтезируется непрерывная цепь, на другой – фрагменты Оказаки (рисунок 26.1, в); 7) ДНК-лигаза – фермент, соединяющий фрагменты Оказаки в единую цепь (рисунок 26.1, г). В результате образуются две идентичные молекулы ДНК (рисунок 26.1, д). 4.4.3. Репарация ДНК - процесс, позволяющий живым организмам восстанавливать повреждения, возникающие в ДНК. Все репарационные механизмы основаны на том, что молекула ДНК состоит из двух полинуклеотидных цепей. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена. Процесс репарации происходит в несколько этапов:
Очень редко происходят повреждения, затрагивающие обе цепи ДНК, т.е. нарушения структуры нуклеотидов комплементарной пары. Такие повреждения в половых клетках не репарируются, так как для осуществления сложной репарации с участием гомологичной рекомбинации требуется наличие диплоидного набора хромосом. Нарушения комплементарности цепей ДНК могут возникать самопроизвольно (спонтанные повреждения) или в результате воздействия физических или химических факторов (индуцируемые повреждения). К спонтанным повреждениям относятся:
К индуцируемым повреждениям относятся повреждения ДНК в результате воздействия радиации или химических веществ:
Ферменты, участвующие в процессе репарации:
|
|
|
Раздел 4.5 |
Транскрипция |
|
|
|
|
4.5.1. Транскрипция – биосинтез РНК на матрице ДНК. Процесс транскрипции также происходит в клеточном ядре. Основные принципы транскрипции:
При участии ДНК-зависимой РНК-полимеразы образуются 3’,5’-фосфодиэфирные связи. Направление полимеризации цепи РНК – от 5’-конца к 3’-концу. Сигналом к окончанию транскрипции служат терминирующие последовательности нуклеотидов (рисунок 8.2,б).
В результате образуются первичные транскрипты – предшественники мРНК, тРНК и рРНК. Затем происходит их посттранскрипционное созревание (процессинг). 4.5.3. Процессинг мРНК включает:
Образующаяся мРНК поступает из ядра в цитоплазму клеток. |
|
| Примеры |
Обучающие задачи и эталоны их решения |
|
|
|
Задачи.1. В молекуле ДНК адениловый нуклеотид составляет 35% от общего содержания нуклеотидов. Рассчитайте (в %) количество каждого из остальных нуклеотидов. 2. В препаратах ДНК, выделенных из двух видов микроорганизмов, содержание тимина составляет соответственно 29 и 16% от общего содержания оснований. Рассчитайте процентное содержание аденина, гуанина и цитозина в этих препаратах ДНК. Какая из ДНК будет более устойчивой к действию высоких температур? Эталоны решения.1. Согласно правилам Чаргаффа, количество аденина в ДНК равно количеству тимина. Поэтому содержание тимидилового нуклеотида также составляет 35%. На долю остальных нуклеотидов приходится 100% - (35 x 2)% = 30%. Количество гуанина в ДНК равняется количеству цитозина, поэтому содержание каждого из них - 30% : 2 = 15%. Итак, А = 35%, Г = 15%, Ц = 15%, Т = 35% (см. 3.4.). 2. Согласно правилам Чаргаффа, содержание оснований в первом препарате ДНК составляет: А = 29%, Г = 21%, Ц = 21%; во втором препарате - А = 16%, Г = 34%, Ц = 34%, очевидно, что содержание пар Г - Ц во втором препарате значительно выше. Поэтому данная молекула ДНК более устойчива к действию температуры (см. 3.4.). |
|
| © С.М.Ершиков, 2007-2012. | |
ОБНОВЛЕНИЯ
08 июня 2019, 16:18:59
08 июня 2019, 16:10:24
07 июня 2019, 22:59:05
09 мая 2019, 16:13:23
27 апреля 2019, 16:00:00
24 апреля 2019, 01:02:18
17 апреля 2019, 22:26:23
17 апреля 2019, 22:21:37
17 апреля 2019, 22:07:42
17 апреля 2019, 22:06:28
ПОДПИСАТЬСЯ НА РАССЫЛКУ
ПРЕДМЕТЫ
- Анатомия
- Акушерство и гинекология
- БЖД, медицина катастроф
- Биохимия
- Биология
- Гистология
- Гигиена
- Генетика
- Диетология
- Дерматовенерология
- Инфекционные болезни
- Культурология
- Лабораторная диагностика
- Летняя практика
- Лучевая диагностика
- Медицинская информатика
- Микробиология
- Неврология
- Общественное здоровье
- Общий уход
О НАС
«Dendrit» - информационный портал для медицинских работников, студентов медицинских ВУЗов, исследователей и пациентов.
Ваш источник новостей и знаний о здоровье.
dendrit.ru © 2014-2024
