Dendrit
Новости, статьи, учебные материалы по медицине
ПОДПИСАТЬСЯ НА РАССЫЛКУ
Ответы 21-40
21. Потребности в углеводах у детей разного возраста. Возрастная характеристика процессов переваривания и всасывания углеводов. Особенности промежуточного обмена углеводов в анте- и постнатальном периоде.
Потребность в углеводах превышает потребность в белках и жирах.
Существенных различий в потребности детей первого года жизни в углеводах при естественном и искусственном вскармливании не отмечается: на протяжении первого года жизни она составляет 12-14 г на 1 кг массы тела.
Очень важно соблюдать правильное соотношение белков, жиров и углеводов. Для детей до 1 года оно составляет 1:3:5 при естественном вскармливании и 1:2:4 при искусственном вскармливании, для детей старше года - 1:1:4.
Основным источником энергии являются углеводы. При сгорании 1 г углеводов высвобождается 4,1 ккал. Жиры удовлетворяют не менее 30% потребностей организма в энергии.
У детей грудного возраста они как источник энергии имеют еще большее значение. Так, у детей первых месяцев жизни, находящихся на грудном вскармливании, за счет жиров покрывается до 50% энергетической потребности, а в первые дни жизни-до 80-90%. При сгорании 1 г жира выделяется 9,0 ккал. Белки также могут использоваться как энергетический материал. При сгорании 1 г белка выделяется 4,0 ккал.
Лактоза, как и другие гликозидазные комплексы, связана с щёточной каемкой и распределена неравномерно по всему тонкому кишечнику. Активность лактазы колеблется в зависимости от возраста. Так, активность лактазы у плода особенно повышена в поздние сроки беременности и сохраняется на высоком уровне до 5-7-летнего возраста. Затем активность фермента снижается, составляя у взрослых 10% от уровня активности, характерного для детей.
22. Переваривание и всасывание углеводов. Катаболизм лактозы и сахарозы. Особенности метаболизма фруктозы и галактозы. Наследственные заболевания углеводного обмена: галактоземия, непереносимость сахарозы и лактозы.
Гидролиз крахмала начинается в ротовой полости. В слюне содержится фермент амилаза, частично расщепляющая крахмал. Основное место переваривания крахмала - тонкий кишечник. Туда поступает амилаза сока поджелудочной железы. Продуктом действия амилазы является мальтоза. Мальтоза далее расщепляется с помощью мальтазы до глюкозы, дисахарид лактоза (содержащаяся в молоке) расщепляется с помощью лактазы до глюкозы и галактозы. Дисахарид сахароза (содержащаяся в пищевом сахаре) расщепляется с помощью сахаразы до глюкозы и фруктозы.
10.5.2. Продукты полного переваривания углеводов - глюкоза, галактоза и фруктоза - через клетки кишечника поступают в кровь путем облегченной диффузии и активного транспорта.
10.5.3. Основные пути метаболизма углеводов в тканях. После всасывания из желудочно-кишечного тракта моносахариды поступают через систему портальной вены в печень. В гепатоцитах происходит превращение галактозы и фруктозы в глюкозу. Таким образом, глюкоза является основным моносахаридом, который поступает в общий кровоток после прохождения углеводов через печень.
Содержание глюкозы в крови здорового человека составляет 3,33 – 5,55 ммоль/л. Глюкоза поглощается из крови клетками всех тканей и органов.
10.5.4. Известны врождённые дефекты ферментов обмена лактозы. Врождённый дефект фермента лактазы приводит к развитию непереносимости лактозы. Употребление в пищу молока сопровождается диспептическими явлениями (рвота, диаррея, метеоризм).
23. Молоко как продукт питания. Основные компоненты женского и коровьего молока. Преимущества грудного вскармливания детей.
(ответа нет)
24. Неорганические метаболиты: натрий, калий, медь, цинк, магний, фтор, йод, селен, сульфат, роль в организме.
Кроме шести главных элементов - С, Н, О, Р, N, S, из которых состоят все органические молекулы, человеку необходимо получать ещё около 20 химических элементов. В зависимости от количества, в каком они должны поступать в организм, минеральные вещества делятся на: макроэлементы - кальций, хлор, магний, калий, натрий - суточная потребность более 100 мг имикроэлементы - железо, марганец, медь, йод, фтор, молибден, селен, цинк и др. - суточная потребность - несколько миллиграммов.
11.6.2. Биологическая роль минеральных веществ: 1. являются структурными компонентами тканей (кальций, фтор); 2. обеспечивают водно-солевой баланс (натрий, калий); 3. являются простетической группой ферментов, входят в состав активных центров, стабилизируют структуру ферментов и фермент-субстратных комплексов (магний, железо, медь); 4. участвуют в передаче нервных импульсов (кальций); 5. участвуют в гормональной регуляции обмена веществ (иод входит в состав гормонов щитовидной железы, цинк – в состав инсулина).
11.6.3. Дефицит микроэлементов в воде и пище может приводить к развитию заболеваний. Например, недостаток железа и меди может вызывать анемию, недостаток фтора способствовать возникновению кариеса, при нехватке йода в пище и воде развивается эндемический зоб.
25. Витамины. Источники витаминов для человека. Причины витаминной недостаточности. Гиповитаминозы, авитаминозы, гипервитаминозы. Витаминсодержащие коферменты. Витаминзависимые и витаминрезистентные состояния у детей.
Витамины - низкомолекулярные органические соединения, поступающие в организм с пищей и обеспечивающие нормальное протекание биохимических и физиологических процессов. Витамины не включаются в структуру тканей и не используются в качестве источника энергии.
11.5.2. Классификация витаминов. Витамины делятся на две группы: витамины, растворимые в воде и витамины, растворимые в жирах. Водорастворимые витамины - В1, В2, В6, В12, РР, Н, С, фолиевая кислота, пантотеновая кислота. Жирорастворимые витамины - А, Д, Е, К.
Для каждого витамина, кроме буквенного обозначения, существует химическое и физиологическое название. Физиологическое название, как правило, состоит из приставки анти- и названия заболевания, развитие которого предупреждает витамин (например, витамин Н - антисеборрейный).
11.5.3. Провитамины. Некоторые витамины могут синтезироваться непосредственно в организме человека. Соединения, служащие предшественниками для синтеза витаминов в клетках организма человека, называются провитаминами. Например, провитамином витамина А является каротин, витамина D2 - эргостерол, D3 - 7-дегидрохолестерол.
11.5.4. Биологическая роль витаминов. Витамины, попадая в организм, превращаются в свою активную форму, которая и принимает непосредственное участие в биохимических процессах.Биологическая роль водорастворимых витаминов заключается в том, что они входят в состав коферментов, участвующих в метаболизме белков, жиров и углеводов в клетках организма человека.
Витамин В1 - тиамин.
Активная форма: кофермент тиаминдифосфат (ТДФ).
Биологическая роль: участвует в реакциях окислительного декарбоксилирования пирувата и α-кетоглутарата.
Суточная потребность: 1-2 мг.
Основные пищевые источники: мука грубого помола, бобовые, мясо, рыба.
Гиповитаминоз: болезнь “бери-бери”. Симптомы: 1. периферические невриты; 2. мышечная слабость; 3. дискоординация движений; 4. увеличение размеров сердца; 5. повышение уровня пирувата в крови. Основная причина смертности у больных бери-бери - сердечная недостаточность.
Витамин В2 - рибофлавин.
Активные формы: коферменты флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД).
Биологическая роль: участвует в окислительно-восстановительных реакциях. Например: 1. перенос электронов в дыхательной и монооксигеназной цепях; 2. окисление сукцината; 3. окисление высших жирных кислот.
Суточная потребность: 1,5 - 3,0 мг.
Основные пищевые источники: молоко, печень, мясо, яйца, жёлтые овощи.
Гиповитаминоз часто встречается у беременных, детей, у людей в состоянии стресса. Симптомы: 1. воспаление сосочков языка -глоссит; 2. растрескивание губ и уголков рта - ангулярный стоматит; 3. помутнение хрусталика - катаракта; 4. воспаление роговицы глаза - кератит.
Витамин В6 - пиридоксин.
Активная форма: кофермент пиридоксальфосфат.
Биологическая роль: - участвует в реакциях: 1. трансаминирования; 2. Декарбоксилирования аминокислот; 3. синтеза никотинамида из триптофана; 4. синтеза δ-аминолевулиновой кислоты (синтез гема).
Суточная потребность: 2 мг.
Основные пищевые источники: хлеб, горох, фасоль, картофель, мясо.
Гиповитаминоз: недостаточность витамина не вызывает специфических симптомов.
Витамин РР - никотинамид (ниацин).
Активная форма: коферменты никотинамидадениндинуклеотид (НАД) и никотинамидадениндинук-леотидфосфат (НАДФ).
Биологическая роль: входит в состав дегидрогеназ. Например: 1. пируватдегидрогеназный комплекс; 2. глюкозо-6-фосфатдегидрогеназа; 3. глутаматдегидрогеназа; 4. β-гидрокси,β-метилглутарил-КоА-редуктаза и многие другие.
Суточная потребность: 15 - 20 мг.
Основные пищевые источники: мясо, рыба,горох, бобы, орехи.
Гиповитаминоз: болезнь пеллагра. Симптомы:
1. дерматит - поражение кожи; 2. диаррея - поражение слизистой желудочно-кишечного тракта; 3. деменция - слабоумие. Поскольку витамин РР может синтезироваться в организме из аминокислоты триптофан, пеллагру можно лечить, вводя в диету дополнительное количество полноценных животных белков. 60 мг триптофана эквивалентны 1 мг никотинамида.
Витамин В9 - фолиевая кислота.
Активная форма: кофермент тетрагидрофолиевая кислота.
Биологическая роль: участвует в реакциях переноса одноуглеродных групп при синтезе: 1. пуриновых нуклеотидов; 2. тимидилового нуклеотида; 3. метионина из гомоцистеина; 4. серина и глицина.
Суточная потребность: 1 - 2,2 мг.
Основные пищевые источники: зелёные листья растений, дрожжи.
Гиповитаминоз: макроцитарная анемия.
Витамин В12 - цианкобаламин.
Активная форма: коферменты метилкобаламин и дезоксиаденозилкобаламин. Имеют сложную структуру, в центре которой находится атом кобальта (Со+), соединённый с четырьмя пиррольными кольцами, образующими корриновоеядро.
Биологическая роль: участвует в реакциях: 1. трансметилирования; 2. обмена серосодержащих аминокислот; 3. образования коферментных форм фолиевой кислоты.
Суточная потребность: 0,003 мг.
Основные пищевые источники: любые продукты животного происхождения.
Гиповитаминоз: мегалобластическая анемия, развивающаяся при нарушении всасывания витамина в кишечнике. Для всасывания витамина В12 в кишечнике, необходим специальный белокгастромукопротеин (транскоррин), получивший название - внутренний фактор Касла.
Этот белок вырабатывается в желудке, связывает витамин В12 (внешний фактор Касла) и образовавшийся комплекс всасывается в кишечнике. Любые причины, приводящие к нарушению выработки желудочного гликопротеина (например, органические поражения желудка, резекция желудка) приводят к гиповитаминозу В12.
Витамин С - аскорбиновая кислота.
Активная форма - неизвестна.
Биологическая роль: кофактор реакций гидроксилирования. Например, в реакциях синтеза: 1. серотонина; 2. оксилизина и оксипролина в коллагене; 3. гомогентизиновой кислоты. Кроме того, способствует поступлению железа в кровь из кишечника и высвобождению его из ферритина. Является антиоксидантом.
Суточная потребность: 50-100 мг.
Гиповитаминоз - болезнь цинга (скорбут). Симптомы: 1. боли в суставах; 2. точечные кровоизлияния - петехии; 3. кровоточивость дёсен; 4. расшатывание зубов; 5. анемия; 6. быстрая утомляемость.
Витамин Н - биотин.
Активная форма: биоцитин.
Биологическая роль - участвует в реакциях карбоксилирования при синтезе: 1. пуриновых нуклеотидов; 2. оксалоацетата; 3. малонил-КоА.
Суточная потребность: 0,26 мг.
Основные пищевые источники: молоко, яичный желток, печень, томаты, шпинат.
Гиповитаминоз: так как витамин синтезируется микрофлорой кишечника, то недостаточность встречается редко. Проявляется в виде специфических дерматитов волосистой части
Болезни нерационального потребления витаминов. Для обеспечения нормального протекания биохимических процессов, в организме человека должен поддерживаться определённый уровень концентрации витаминов. При изменении этого уровня развиваются заболевания с симптомами, харктерными для каждого витамина.
Гипервитаминозы - заболевания, вызванные избыточным содержанием витаминов в организме. Характерны для жирорастворимых витаминов, способных накапливаться в клетках печени. Чаще всего встречаются гипервитаминозы А и D, связанные с передозировкой их лекарственных препаратов. Гипервитаминоз А характеризуется общими симптомами отравления: сильными головными болями, тошнотой, слабостью. Гипервитаминоз D сопровождается деминерализацией костей, кальцинацией мягких тканей, образованием камней в почках.
Гиповитаминозы - заболевания, вызванные недостатком витаминов в организме. Первичные гиповитаминозы связаны с нарушением процессов поступления витаминов в организм при: 1. недостатке витаминов в пище; 2. ускоренном распаде витаминов в кишечнике под действием патогенной микрофлоры; 3. нарушении синтеза витаминов кишечной микрофлорой при дисбактериозе; 4. нарушении всасывания витаминов; 5. приеме лекарственных препаратов - антивитаминов. Вторичные гиповитаминозы связаны с нарушением процессов превращения витаминов в их активные формы в клетках организма человека. Причиной могут служить генетические дефекты или нарушения биохимических процессов при различных заболеваниях органов и тканей.
Авитаминозы - заболевания, вызванные полным отсутствием витамина в организме.
26. Жирорастворимые витамины А, Е и К – биологическая роль, пищевые источники, причины и проявления гипо- и гипервитаминоза.
Витамин А - ретинол.
Активная форма: цис-ретиналь.
Биологическая роль: 1. контролирует рост и дифференцировку быстро пролиферирующих тканей (эмбриональной, хрящевой, костной, эпителиальной); 2. участвует в фотохимическом акте зрения.
Суточная потребность: 0,5-2,0 мг.
Основные пищевые источники: сливочное масло, печень животных и рыб, b-каротин в красных плодах.
Участие витамина А в механизме сумеречного зрения. В процессе светоощущения главная роль принадлежит пигменту родопсину - сложному белку, состоящему из белка опсина и простетическойгруппы - цис-ретиналя. Под действием света цис-ретиналь света превращается в изомер - транс-ретиналь, что приводит к разрушению пигмента родопсина и возникновению нервного импульса. Восстановление пигмента происходит по схеме:
Процесс изомеризации транс-ретинола в сетчатке глаза протекает очень медленно. Основное его количество поступает в кровь, затем в печень, где и происходит быстрое превращение транс-ретинола в цис-ретинол, который попадает в кровь и поглощается сетчаткой глаза. Процесс лимитируется запасом в печени транс-ретинола (витамина А).
Гиповитаминоз: нарушение темновой адаптации зрения (ночная слепота) у взрослых; у детей - остановка роста, ороговение эпителия всех органов - гиперкератоз, сухость роговицы глаза - ксерофтальмия, размягчение роговицы под действием микрофлоры - кератомаляция.
Витамин К - филлохинон.
Активная форма: неизвестна.
Биологическая роль – участие в синтезе белковых факторов свёртывания крови: II (протромбин), VII (проконвертин), IX (Кристмас-фактор), и X (фактор Прауэра-Стюарта).
Суточная потребность: 1 мг.
Основные пищевые источники: синтезируется микрофлорой кишечника.
Гиповитаминоз - паренхиматозные и капиллярные кровотечения.
Витамин Е - токоферол.
Активная форма: неизвестна.
Биологическая роль - природный антиоксидант, тормозит пероксидное окисление липидов клеточных мембран.
Суточная потребность: 5 мг.
Основные пищевые источники: растительные масла.
Гиповитаминоз у человека - гемолитическая анемия. У животных - мышечная дистрофия, дегенерация спинного мозга, атрофия семенников, пероксидный гемолиз эритроцитов.
27. Витамин D. Образование активной формы витамина из провитамина. Биологическая роль. Нарушения обмена при недостаточности витамина D3 у детей.
Витамин D3 - холекальциферол.
Активная форма: 1,25-дигидроксихолекальциферол, кальцитриол.
Биологическая роль: 1. регуляция всасывания ионов кальция и фосфатов в кишечнике; 2. реабсорбция ионов кальция в почечных канальцах; 3. мобилизация ионов кальция из костей.
Суточная потребность: 10-15 мкг (500-1000 МЕ).
Основные пищевые источники: печень животных и рыб, яйца, молоко, сливочное масло.
Синтез витамина D3 и его активных форм в тканях человека. Предшественником (провитамином) витамина D3 в организме человека служит 7-дегидрохолестерол, который при действиина кожу ультрафиолетового излучения, переходит в холекальциферол.
Образование активной формы витамина происходит последовательно в печени и почках путём гидроксилирования по 1 и 25 углеродным атомам. Образующийся 1,25-дигидроксихолекальциферол обладает гормональной активностью (кальцитриол). Тканями-мишенями для него являются кишечник, почки, кости. В эпителии кишечника и почечных канальцах кальцитриол индуцирует синтез Са-связывающего белка, что способствует всасыванию ионов Са2+ из пищи и реабсорбции их почками. В костной ткани угнетает синтез коллагена, уменьшает Са- связывающую способность, что приводит к мобилизации кальция из костей.
Гиповитаминоз: у детей - рахит. Симптомы: 1. снижение мышечного тонуса; 2. деформация костей черепа, груди, позвоночника, нижних конечностей. У взрослых - остеопороз - деминерализация костей.
При нарушении образования активных форм витамина D3 (например, при поражении печени и почек) развивается D-резистентное рахитоподобное состояние.
28. Витамин B1 — коферментные функции, биологическая роль, суточная потребность, пищевые источники, строение, нарушения обмена при недостаточности тиамина.
Витамин В1 - тиамин.
Активная форма: кофермент тиаминдифосфат (ТДФ).
Биологическая роль: участвует в реакциях окислительного декарбоксилирования пирувата и α-кетоглутарата.
Суточная потребность: 1-2 мг.
Основные пищевые источники: мука грубого помола, бобовые, мясо, рыба.
Гиповитаминоз: болезнь “бери-бери”. Симптомы: 1. периферические невриты; 2. мышечная слабость; 3. дискоординация движений; 4. увеличение размеров сердца; 5. повышение уровня пирувата в крови. Основная причина смертности у больных бери-бери - сердечная недостаточность.
29. Витамин B2 — коферментные функции, биологическая роль, суточная потребность, пищевые источники, строение.
Витамин В2 - рибофлавин.
Активные формы: коферменты флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД).
Биологическая роль: участвует в окислительно-восстановительных реакциях. Например: 1. перенос электронов в дыхательной и монооксигеназной цепях; 2. окисление сукцината; 3. окисление высших жирных кислот.
Суточная потребность: 1,5 - 3,0 мг.
Основные пищевые источники: молоко, печень, мясо, яйца, жёлтые овощи.
Гиповитаминоз часто встречается у беременных, детей, у людей в состоянии стресса. Симптомы: 1. воспаление сосочков языка -глоссит; 2. растрескивание губ и уголков рта - ангулярный стоматит; 3. помутнение хрусталика - катаракта; 4. воспаление роговицы глаза - кератит.
30. Витамин РР — коферментные функции, биологическая роль, суточная потребность, пищевые источники, строение.
Витамин РР - никотинамид (ниацин).
Активная форма: коферменты никотинамидадениндинуклеотид (НАД) и никотинамидадениндинук-леотидфосфат (НАДФ).
Биологическая роль: входит в состав дегидрогеназ. Например: 1. пируватдегидрогеназный комплекс; 2. глюкозо-6-фосфатдегидрогеназа; 3. глутаматдегидрогеназа; 4. β-гидрокси,β-метилглутарил-КоА-редуктаза и многие другие.
Суточная потребность: 15 - 20 мг.
Основные пищевые источники: мясо, рыба,горох, бобы, орехи.
Гиповитаминоз: болезнь пеллагра. Симптомы:
1. дерматит - поражение кожи; 2. диаррея - поражение слизистой желудочно-кишечного тракта; 3. деменция - слабоумие. Поскольку витамин РР может синтезироваться в организме из аминокислоты триптофан, пеллагру можно лечить, вводя в диету дополнительное количество полноценных животных белков. 60 мг триптофана эквивалентны 1 мг никотинамида.
31. Витамин B6 —коферментные функции, биологическая роль, пищевые источники, строение. Потребность в витамине B6 в зависимости от качественного состава пищевого рациона.
Витамин В6 - пиридоксин.
Активная форма: кофермент пиридоксальфосфат.
Биологическая роль: - участвует в реакциях: 1. трансаминирования; 2. Декарбоксилирования аминокислот; 3. синтеза никотинамида из триптофана; 4. синтеза δ-аминолевулиновой кислоты (синтез гема).
Суточная потребность: 2 мг.
Основные пищевые источники: хлеб, горох, фасоль, картофель, мясо.
Гиповитаминоз: недостаточность витамина не вызывает специфических симптомов.
32. Фолиевая кислота - строение, коферментные функции, биологическая роль, суточная потребность, источники. Проявления недостаточности фолиевой кислоты. Механизм бактериостатического действия сульфаниламидных препаратов.
форма: кофермент тетрагидрофолиевая кислота.
Биологическая роль: участвует в реакциях переноса одноуглеродных групп при синтезе: 1. пуриновых нуклеотидов; 2. тимидилового нуклеотида; 3. метионина из гомоцистеина; 4. серина и глицина.
Суточная потребность: 1 - 2,2 мг.
Основные пищевые источники: зелёные листья растений, дрожжи.
Гиповитаминоз: макроцитарная анемия.
33. Витамин B12 – биологическая роль, суточная потребность, источники. Причины недостаточности витамина B12 в организме и ее проявления. Роль "внутреннего фактора Касла" в усвоении витамина B12.
Витамин В12 - цианкобаламин.
Активная форма: коферменты метилкобаламин и дезоксиаденозилкобаламин. Имеют сложную структуру, в центре которой находится атом кобальта (Со+), соединённый с четырьмя пиррольными кольцами, образующими корриновоеядро.
Биологическая роль: участвует в реакциях: 1. трансметилирования; 2. обмена серосодержащих аминокислот; 3. образования коферментных форм фолиевой кислоты.
Суточная потребность: 0,003 мг.
Основные пищевые источники: любые продукты животного происхождения.
Гиповитаминоз: мегалобластическая анемия, развивающаяся при нарушении всасывания витамина в кишечнике. Для всасывания витамина В12 в кишечнике, необходим специальный белокгастромукопротеин (транскоррин), получивший название - внутренний фактор Касла.
Этот белок вырабатывается в желудке, связывает витамин В12 (внешний фактор Касла) и образовавшийся комплекс всасывается в кишечнике. Любые причины, приводящие к нарушению выработки желудочного гликопротеина (например, органические поражения желудка, резекция желудка) приводят к гиповитаминозу В12.
34. Витамин С – биологическая роль, суточная потребность, пищевые источники, строение, нарушения обмена при недостаточности аскорбиновой кислоты.
Витамин С - аскорбиновая кислота.
Активная форма - неизвестна.
Биологическая роль: кофактор реакций гидроксилирования. Например, в реакциях синтеза: 1. серотонина; 2. оксилизина и оксипролина в коллагене; 3. гомогентизиновой кислоты. Кроме того, способствует поступлению железа в кровь из кишечника и высвобождению его из ферритина. Является антиоксидантом.
Суточная потребность: 50-100 мг.
Гиповитаминоз - болезнь цинга (скорбут). Симптомы: 1. боли в суставах; 2. точечные кровоизлияния - петехии; 3. кровоточивость дёсен; 4. расшатывание зубов; 5. анемия; 6. быстрая утомляемость.
35. Безопасность пищи. Химические и биологические загрязнители, их влияние на обмен веществ. Метаболизм этанола.
Важнейшая функция печени - детоксикационная (или барьерная). Она имеет существенное значение для сохранения жизни организма. В печени происходит обезвреживание таких веществ, как билирубин и продукты катаболизма аминокислот в кишечнике, а также инактивируются лекарственные препараты и токсические вещества экзогенного происхождения, NH3 - продукт азотистого обмена, который в результате ферментативных реакций превращается в нетоксичную мочевину, гормоны и биогенные амины.
Вещества, поступающие в организм из окружающей среды и не используемые им для построения тканей организма или как источники энергии, называют чужеродными веществами, или ксенобиотиками. Эти вещества могут попадать в организм с пищей, через кожу или с вдыхаемым воздухом.
Чужеродные вещества, или ксенобиотики, делят на 2 группы:
- продукты хозяйственной деятельности человека (промышленность, сельское хозяйство, транспорт);
- вещества бытовой химии - моющие средства, вещества для борьбы с насекомыми, парфюмерия.
Гидрофильные ксенобиотики выводятся из организма в неизменённом виде с мочой, гидрофобные могут задерживаться в тканях, связываясь с белками или образуя комплексы с липидами клеточных мембран. Со временем накопление в клетках тканей чужеродного вещества приведёт к нарушению их функций. Для удаления таких ненужных для организма веществ в процессе эволюции выработались механизмы их детоксикации (обезвреживания) и выведения из организма.
Катаболизм этилового спирта осуществляется главным образом в печени. Здесь окисляется от 75% до 98% введённого в организм этанола.
Окисление алкоголя - сложный биохимический процесс, в который вовлекаются основные метаболические процессы клетки. Превращение этанола в печени осуществляется тремя путями с образованием токсического метаболита – ацетальдегида.
Основную роль в метаболизме этанола играет цинксодержащий NAD+- зависимый фермент - алкогольдегидрогеназа, локализующаяся в основном в цитозоле и митохондриях печени (95%). В ходе реакции происходит дегидрирование этанола, образуются ацетальдегид и восстановленный кофермент NADH.
Алкогольдегидрогеназа катализирует обратимую реакцию, направление которой зависит от концентрации ацетальдегида и соотношения NADH/NAD+ в клетке.
С2Н5ОН + HAD+ ↔ СН3СНО + NADH + H+.
Фермент алкогольдегидрогеназа - димер, состоящий из идентичных или близких по первичной структуре полипептидных цепей, кодируемых аллелями одного гена. Существуют 3 изоформы алкогольдегидрогеназы (АДГ): АДГ1, АДГ2, АДГ3, различающиеся по строению протомеров, локализации и активности. Для европейцев характерно присутствие изоформ АДГ1 и АДГ3. У некоторых восточных народов преобладает изоформа АДГ2, характеризующаяся высокой активностью, это может быть причиной их повышенной чувствительности к алкоголю. При хроническом алкоголизме количество фермента в печени не увеличивается, т.е. он не является индуцируемым ферментом.
Б. Окисление этанола при участии цитохром Р450 - зависимой микросомальной этанолокисляющей системы системы
Цитохром Р450-зависимая микросомальная этанолокисляющая сисгема (МЭОС) локализована в мембране гладкого ЭР гепатоцитов. МЭОС играет незначительную роль в метаболизме небольших количеств алкоголя, но индуцируется этанолом, другими спиртами, лекарствами типа барбитуратов и приобретает существенное значение при злоупотреблении этими веществами. Этот путь окисления этанола происходит при участии одной из изоформ Р450 - изофермента Р450 II E1. При хроническом алкоголизме окисление этанола ускоряется на 50 - 70% за счёт гипертрофии ЭР и индукции цитохрома Р450 II E1.
С2Н5ОН + NADPH + Н+ + О2 → СН3СНО + NADP+ + 2Н2О.
Кроме основной реакции, цитохром Р450 катализирует образование активных форм кислорода (О2-, Н2О2), которые стимулируют ПОЛ в печени и других органах (см. раздел 8).
В. Окисление этанола каталазой
Второстепенную роль в окислении этанола играет каталаза, находящаяся в пероксисомах цитоплазмы и митохондрий клеток печени. Этот фермент расщепляет примерно 2% этанола, но при этом утилизирует пероксид водорода.
СН3СН2ОН + Н2О2 → СН3СНО + 2Н2О.
Г. Метаболизм и токсичность ацетальдегида
Ацетальдегид, образовавшийся из этанола, окисляется до уксусной кислоты двумя ферментами: FAD -зависимой альдегидоксидазой и NAD+ -зависимой ацетальдегиддегидрогеназой (АлДГ).
СН3СНО + О2 + H2O → СН3СООН + Н2О2 .
Повышение концентрации ацетальдегида в клетке вызывает индукцию фермента алъдегидоксидазы. В ходе реакции образуются уксусная кислота, пероксид водорода и другие активные формы кислорода, что приводит к активации ПОЛ.
Другой фермент ацетальдегиддегидрогеназа (АлДГ) окисляет субстрат при участии кофермента NAD+.
СН3СНО + Н2О + NAD+ → СН3СООН + NADH + H+.
Полученная в ходе реакции уксусная кислота активируется под действием фермента ацетил-КоА-синтетазы. Реакция протекает с использованием кофермента А и молекулы АТФ. Образовавшийся ацетил-КоА, в зависимости от соотношения АТФ/АДФ и концентрации окса-лоацетата в митохондриях гепатоцитов, может "сгорать" в ЦТК, идти на синтез жирных кислот или кетоновых тел.
В разных тканях организма человека встречаются полиморфные варианты АлДГ. Они характеризуются широкой субстратной специфичностью, разным распределением по клеткам тканей (почки, эпителий, слизистая оболочка желудка и кишечника) и в компартментах клетки. Например, изоформа АлДГ, локализованная в митохондриях гепатоцитов, обладает более высоким сродством к ацетальдегиду, чем цитозольная форма фермента.
Ферменты, участвующие в окислении этанола, - алкогольдегидрогеназа и АлДГ по разному распределены: в цитозоле - 80%/20% и митохондриях - 20%/80%. При поступлении больших доз алкоголя (более 2 г/кг) из-за разных скоростей окисления этанола и ацетальдегида в цитозоле резко повышается концентрация последнего. Ацетальдегид - очень реакционно-способное соединение; он неферментативно может ацетилировать SH-, NH2- группа белков и других соединений в клетке и нарушать их функции. В модифицированных (ацетилированных) белках могут возникать "сшивки", нехарактерные для нативной структуры (например, в белках межклеточного матрикса - эластине и коллагене, некоторых белках хроматина и липопротеинов, формирующихся в печени). Ацетилирование ядерных, цитоплазматических ферментов и структурных елков приводит к снижению синтеза экспортируемых печенью в кровь белков, например альбумина, который, удерживая Na+, поддерживает коллоидно-осмотическое давление, а также участвует в транспорте многих гидрофобных веществ в крови (см. раздел 14). Нарушение функций альбумина в сочетании с повреждающим действием ацетальдегида на мембраны сопровождается поступлением в клетки по градиенту концентрации ионов натрия и воды, происходит осмотическое набухание этих клеток и нарушение их функций.
Активное окисление этанола и ацетальдегида приводит к увеличению отношения NADH/NAD+, что снижает активность NAD+-зависимых ферментов в цитозоле и менее значительно в митохондриях.
36. Катаболические и анаболические пути обмена. Функции метаболизма. Три стадии катаболизма основных питательных веществ в организме. Связь общего пути катаболизма с цепью переноса электронов и протонов и синтезом АТФ. Роль НАД- и ФАД-зависимых дегидрогеназ.
Метаболизм (обмен веществ) – совокупность химических реакций, протекающих в живой клетке. Эти реакции протекают в определённой последовательности и тесно связаны между собой. Главные функции метаболизма в клетке:
а) запасание энергии, которая добывается путем расщепления пищевых веществ, поступающих в организм, или путем преобразования энергии солнечного света;
б) превращение молекул пищевых веществ в строительные блоки;
в) сборку белков, нуклеиновых кислот, липидов, полисахаридов и прочих клеточных компонентов из этих строительных блоков;
г) синтез и разрушение тех биомолекул, которые необходимы для выполнения специфических функций данной клетки.
13.1.2. Метаболизм включает множество метаболических путей. Метаболический путь - определенная последовательность ферментативных реакций в клетке. Промежуточные продукты реакций метаболического пути называются метаболитами. На каждой из стадий метаболического пути происходит небольшое химическое изменение метаболитов. В результате этих превращений исходная молекула превращается в конечный продукт метаболического пути.
13.1.3. Большинство метаболических путей являются линейными, но встречаются и циклические метаболические пути. Обычно метаболические пути имеют разветвления, в которых одни вещества выходят из цепи реакций данного пути, а другие, наоборот, включаются в него.
Обратите внимание, что главные метаболические пути немногочисленны и одинаковы у большинства живых организмов. Именно эти пути представляют для нас наибольший интерес. Рассмотрению их будут посвящены разделы 13.3. и 13.4 настоящей темы.
13.1.4. Две стороны (фазы) метаболизма. Метаболизм складывается из двух противоположных сторон : катаболизма и анаболизма.
Катаболизм – это фаза, в которой происходит последовательное расщепление сложных молекул до более простых, таких, как СО2, вода и аммиак. Процессы катаболизма сопровождаются выделением энергии. Эта энергия частично аккумулируется в форме макроэргического соединения – аденозинтрифосфата (АТФ).
Анаболизм – это фаза метаболизма, в которой происходит образование (биосинтез) сложных молекул (белков, липидов, полисахаридов) из простых предшественников. Процессы биосинтеза протекают с затратой энергии. Источником этой энергии служит распад АТФ до АДФ и неорганического фосфата.
Метаболические пути, выполняющие как катаболическую, так и анаболическую функцию, называют амфиболическими.
13.1.5. Катаболическая и анаболическая фазы метаболизма тесно связаны между собой (рисунок 13.2):
а) Энергия, выделяемая в реакциях катаболизма, и аккумулированная в форме молекул АТФ, потребляется в анаболических процессах.
б) В реакциях катаболизма образуются простые метаболиты, которые могут использоваться в реакциях биосинтеза (анаболизма).
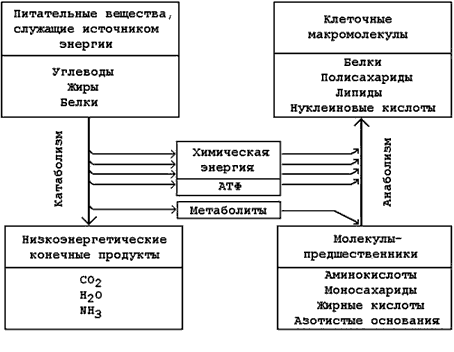
Рисунок 13.2. Взаимосвязь путей катаболизма и анаболизма
Как уже было сказано (раздел 13.1), катаболизм – совокупность химических реакций превращения высокомолекулярных соединений в низкомолекулярные. Это сложный ферментативный процесс, в котором принято выделять три основные стадии (рисунок 13.3). Для каждой стадии запомните, какие вещества вступают и образуются, локализацию в организме и энергетическую значимость.
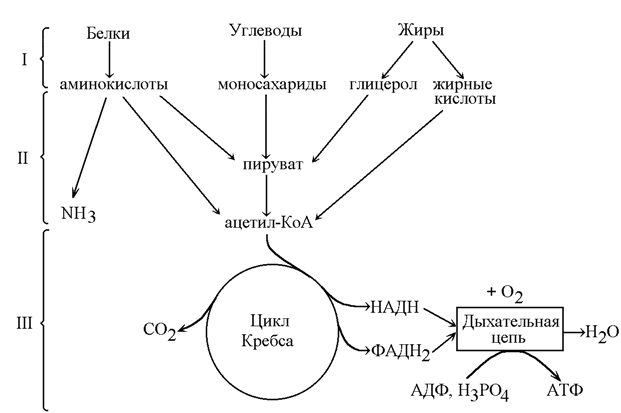
Рисунок 13.3. Три стадии катаболизма питательных веществ (обозначены римскими цифрами).
13.2.2. На первой стадии крупные биомолекулы расщепляются на составляющие их строительные блоки: полисахариды превращаются в пентозы и гексозы, жиры – в жирные кислоты, глицерол и другие компоненты, белки – в аминокислоты. Это происходит в желудочно-кишечном тракте, а также в лизосомах клетки. Реакции катализируют ферменты, относящиеся к классу гидролаз. Относительная энергоотдача составляет менее 1% всей высвобождаемой энергии.
На второй стадии строительные блоки превращаются в более простые молекулы. Моносахариды, глицерол и большинство аминокислот расщепляются до одного и того же трёхуглеродного метаболита – пирувата. Это происходит в цитоплазме клеток. В дальнейшем пируват, а также жирные кислоты и некоторые аминокислоты окисляются до ацетильного остатка, связанного с коэнзимом А (ацетил-КоА). Эти реакции протекают уже в митохондриях клетки. Пируват и ацетил-КоА, находящиеся на пересечении нескольких метаболических путей, можно отнести кключевым или узловым метаболитам. Относительная энергоотдача второй стадии катаболизма около 20%; выделяемая энергия может быть частично аккумулирована в виде АТФ.
На третьей стадии происходит окисление ацетильной группы в цикле трикарбоновых кислот Кребса до СО2 и восстановленных форм коферментов НАД и ФАД. Эти коферменты окисляются в дыхательной цепи до Н2О; выделяемая энергия аккумулируется в АТФ. Все эти реакции протекают в митохондриях. Относительная энергоотдача третьей стадии - около 80%.
13.2.3. Различают общий и специфические пути катаболизма. К специфическим путям катаболизма веществ того или иного класса относят главным образом реакции первой и второй стадии катаболизма, которые для каждого класса могут существенно различаться. Третья стадия, одинаковая для всех классов питательных веществ, называется общим путём катаболизма.
37. Пировиноградная кислота: пути образования и использования в организме. Окислительное декарбоксилирование пировиноградной кислоты. Строение пируватдегидрогеназного комплекса. Значение витаминных коферментов в декарбоксилировании пирувата.
Заключительной реакцией второй стадии катаболизма питательных веществ является окислительное декарбоксилирование пирувата.
Пируват подвергается окислительному декарбоксилированию до ацетил-коэнзима А (ацетил-КоА) в митохондриях клеток при участии пируватдегидрогеназного комплекса. Пируватдегидрогеназный комплекс – мультифермент, в состав которого входит 3 фермента:
1) пируватдекарбоксилаза (коферментом служит производное витамина В1 - тиаминдифосфат, ТДФ, формулу его см. в теме «Коферменты»);
2) трансацилаза (кофермент – липоевая кислота, ЛК
3) дигидролипоилдегидрогеназа (в качестве кофермента используется производное витамина В2 – флавинадениндинуклеотид, ФАД, формулу его см. в теме «Коферменты»).
В реакции участвуют также два кофермента: коэнзим А (НSКоА, производное витамина В3) и никотинамидадениндинуклеотид (НАД, производное витамина РР), связанные при помощи нековалентных связей соответственно с трансацилазой и дигидролипоилдегидрогеназой.
Ацетил-КоА затем подвергается окислению в цикле Кребса, а НАДН служит донором водорода для дыхательной цепи. Процесс является аэробным, т.к. конечный акцептор водорода НАДН – кислород. Скорость реакции снижается при накоплении в клетке ацетил-КоА, НАДН и АТФ, увеличивается – при возрастании концентрации АДФ. Регуляторным ферментом комплекса является пируватдекарбоксилаза.
Аналогично происходит окислительное декарбоксилирование α-кетоглутарата – одного из метаболитов цикла Кребса. Реакцию катализирует α-кетоглутаратдегидрогеназный комплекс, сходный с пируватдегидрогеназным комплексом.
38. Ацетил-КоА: источники и основные пути использования в тканях. Компартментализация обмена ацетил-КоА.
Дальнейшем пируват, а также жирные кислоты и некоторые аминокислоты окисляются до ацетильного остатка, связанного с коэнзимом А (ацетил-КоА). Эти реакции протекают уже в митохондриях клетки. Пируват и ацетил-КоА, находящиеся на пересечении нескольких метаболических путей, можно отнести кключевым или узловым метаболитам.
39. Цикл трикарбоновых кислот: последовательность реакций, характеристика ферментов. Амфиболическая функция цитратного цикла. Связь с обменом углеводов, жиров и белков.
Реакции цикла Кребса относятся к третьей стадии катаболизма питательных веществ и происходят в митохондриях клетки. Эти реакции относятся к общему пути катаболизма и характерны для распада всех классов питательных веществ (белков, липидов и углеводов).
Главной функцией цикла является окисление ацетильного остатка с образованием четырёх молекул восстановленных коферментов (трёх молекул НАДН и одной молекулы ФАДН2), а также образование молекулы ГТФ путём субстратного фосфорилирования. Атомы углерода ацетильного остатка выделяются в виде двух молекул СО2.
13.4.2. Цикл Кребса включает 8 последовательных стадий, обращая особое внимание на реакции дегидрирования субстратов:
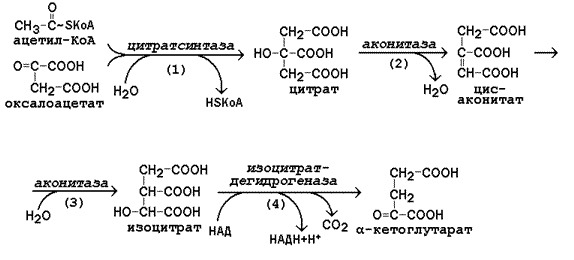
Рисунок 13.6. Реакции цикла Кребса, включая образование α-кетоглутарата
а) конденсация ацетил-КоА с оксалоацетатом, в результате которой образуется цитрат (рис.13.6, реакция 1); поэтому цикл Кребса называют также цитратным циклом. В этой реакции метильный углерод ацетильной группы взаимодействует с кетогруппой оксалоацетата; одновременно происходит расщепление тиоэфирной связи. В реакции освобождается КоА-SH, который может принять участие в окислительном декарбоксилировании следующей молекулы пирувата. Реакцию катализирует цитратсинтаза, это – регуляторный фермент, он ингибируется высокими концентрациями НАДН, сукцинил-КоА, цитрата.
б) превращение цитрата в изоцитрат через промежуточное образование цис-аконитата. Образующийся в первой реакции цикла цитрат содержит третичную гидроксильную группу и не способен окисляться в условиях клетки. Под действием фермента аконитазы идёт отщепление молекулы воды (дегидратация), а затем её присоединение (гидратация), но другим способом (рис.13.6, реакции 2-3). В результате данных превращений гидроксильная группа перемещается в положение, благоприятствующее её последующему окислению.
в) дегидрирование изоцитрата с последующим выделением молекулы СО2 (декарбоксилированием) и образованием α-кетоглутарата (рис. 13.6, реакция 4). Это – первая окислительно-восстановительная реакция в цикле Кребса, в результате которой образуется НАДН. Изоцитратдегидрогеназа, катализирующая реакцию, - регуляторный фермент, активируется АДФ. Избыток НАДН ингибирует фермент.
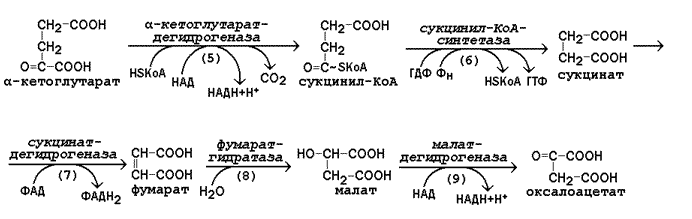
Рисунок 13.7. Реакции цикла Кребса, начиная с α-кетоглутарата.
г) окислительное декарбоксилирование α-кетоглутарата, катализируется мультиферментным комплексом (рис. 13.7, реакция 5), сопровождается выделением СО2 и образованием второй молекулы НАДН. Эта реакция аналогична пируватдегидрогеназной реакции. Ингибитором служит продукт реакции – сукцинил-КоА.
д) субстратное фосфорилирование на уровне сукцинил-КоА, в ходе которого энергия, освобождающаяся при гидролизе тиоэфирной связи, запасается в форме молекулы ГТФ. В отличие от окислительного фосфорилирования, этот процесс протекает без образования электрохимического потенциала митохондриальной мембраны (рис. 13.7, реакция 6).
е) дегидрирование сукцината с образованием фумарата и молекулы ФАДН2 (рис. 13.7, реакция 7). Фермент сукцинатдегидрогеназа прочно связан с внутренней мембраной митохондрии.
ж) гидратация фумарата, в результате чего в молекуле продукта реакции появляется легко окисляемая гидроксильная группа (рис. 13.7, реакция 8).
з) дегидрирование малата, приводящее к образованию оксалоацетата и третьей молекулы НАДН (рис.13.7, реакция 9). Образующийся в реакции оксалоацетат может вновь использоваться в реакции конденсации с очередной молекулой ацетил-КоА (рис. 13.6, реакция 1). Поэтому данный процесс носит циклический характер.
13.4.3. Таким образом, в результате описанных реакций подвергается полному окислению ацетильный остаток СН3-СО-. Количество молекул ацетил-КоА, превращаемых в митохондриях в единицу времени, зависит от концентрации оксалоацетата. Основные пути увеличения концентрации оксалоацетата в митохондриях (соответствующие реакции будут рассмотрены позднее):
а) карбоксилирование пирувата – присоединение к пирувату молекулы СО2 с затратой энергии АТФ;
б) дезаминирование или трансаминирование аспартата – отщепление аминогруппы с образованием на её месте кетогруппы.
13.4.4. Некоторые метаболиты цикла Кребса могут использоваться для синтеза структурных блоков для построения сложных молекул. Так, оксалоацетат может превращаться в аминокислоту аспартат, а α–кетоглутарат – в аминокислоту глутамат. Сукцинил-КоА принимает участие в синтезе гема – простетической группы гемоглобина. Таким образом, реакции цикла Кребса могут участвовать как в процессах катаболизма, так и анаболизма, то есть цикл Кребса выполняет амфиболическую функцию (см. 13.1).
40. Окисление НАДН2 и ФАДН2 в митохондриях. Характеристика основных компонентов дыхательной цепи. Ферментные комплексы. Дегидрирование субстратов и окисление водорода как источник энергии для синтеза АТФ. Роль АТФ в организме.
В пируватдегидрогеназной реакции и в цикле Кребса происходит дегидрирование (окисление) субстратов (пируват, изоцитрат, α-кетоглутарат, сукцинат, малат). В результате этих реакций образуются НАДН и ФАДН2. Эти восстановленные формы коферментов окисляются в митохондриальной дыхательной цепи. Окисление НАДН и ФАДН2, протекающее сопряжённо с синтезом АТФ из АДФ и Н3РО4 называется окислительным фосфорилированием.
Схема строения митохондрии показана на рисунке 14.1. Митохондрии представляют собой внутриклеточные органеллы, имеющие две мембраны: наружную (1) и внутреннюю (2). Внутренняя митохондриальная мембрана образует многочисленные складки – кристы (3). Пространство, ограниченное внутренней митохондриальной мембраной, носит название матрикс (4), пространство, ограниченное наружной и внутренней мембранами, - межмембранное пространство
Дыхательная цепь – последовательная цепь ферментов, осуществляющая перенос ионов водорода и электронов от окисляемых субстратов к молекулярному кислороду – конечному акцептору водорода. В ходе этих реакций выделение энергии происходит постепенно, небольшими порциями, и она может быть аккумулирована в форме АТФ. Локализация ферментов дыхательной цепи – внутренняя митохондриальная мембрана.
Дыхательная цепь включает четыре мультиферментных комплекса
I. НАДН-KoQ-редуктаза (содержит промежуточные акцепторы водорода: флавинмононуклеотид и железосерные белки).
II. Сукцинат-KoQ-редуктаза (содержит промежуточные акцепторы водорода: ФАД и железосерные белки).
III. KoQН2-цитохром с-редуктаза (содержит акцепторы электронов: цитохромы b и с1, железосерные белки).
IV. Цитохром с-оксидаза (содержит акцепторы электронов: цитохромы а и а3, ионы меди Cu2+).
14.1.3. В качестве промежуточных переносчиков электронов выступают убихинон (коэнзим Q) и цитохром с.
Убихинон (KoQ) – жирорастворимое витаминоподобное вещество, способен легко диффундировать в гидрофобной фазе внутренней мембраны митохондрий. Биологическая роль коэнзима Q – перенос электронов в дыхательной цепи от флавопротеинов (комплексы I и II) к цитохромам (комплекс III).
Цитохром с – сложный белок, хромопротеин, простетическая группа которого – гем – содержит железо с переменной валентностью (Fe3+ в окисленной форме и Fe2+ в восстановленной форме). Цитохром с является водорастворимым соединением и располагается на периферии внутренней митохондриальной мембраны в гидрофильной фазе. Биологическая роль цитохрома с – перенос электронов в дыхательной цепи от комплекса III к комплексу IV.
14.1.4. Промежуточные переносчики электронов в дыхательной цепи расположены в соответствии с их окислительно-восстановительными потенциалами. В этой последовательности способность отдавать электроны (окисляться) убывает, а способность присоединять электроны (восстанавливаться) возрастает. Наибольшей способности отдавать электроны обладает НАДН, наибольшей способностью присоединять электроны – молекулярный кислород.
Механизм синтеза АТФ описывает хемиосмотическая теория (автор - П. Митчелл). Согласно этой теории, компоненты дыхательной цепи, расположенные во внутренней митохондриальной мембране, в ходе переноса электронов могут «захватывать» протоны из матрикса митохондрий и передавать их в межмембранное пространство. При этом наружная поверхность внутренней мембраны приобретает положительный заряд, а внутренняя – отрицательный, т.е. создаётся градиент концентрации протонов с более кислым значением рН снаружи. Так возникает трансмембранный потенциал (ΔµН+). Существует три участка дыхательной цепи, на которых он образуется. Эти участки соответствуют I, III и IV комплексам цепи переноса электронов.
Энергия, аккумулированная в форме АТФ, используется в организме для обеспечения разнообразных биохимических и физиологических процессов. Запомните основные примеры использования энергии АТФ:
1) синтез сложных химических веществ из более простых (реакции анаболизма);
2) сокращение мышц (механическая работа);
3) образование трансмембранных биопотенциалов;
4) активный транспорт веществ через биологические мембраны.
ОБНОВЛЕНИЯ
08 июня 2019, 16:18:59
08 июня 2019, 16:10:24
07 июня 2019, 22:59:05
09 мая 2019, 16:13:23
27 апреля 2019, 16:00:00
24 апреля 2019, 01:02:18
17 апреля 2019, 22:26:23
17 апреля 2019, 22:21:37
17 апреля 2019, 22:07:42
17 апреля 2019, 22:06:28
ПОДПИСАТЬСЯ НА РАССЫЛКУ
ПРЕДМЕТЫ
- Анатомия
- Акушерство и гинекология
- БЖД, медицина катастроф
- Биохимия
- Биология
- Гистология
- Гигиена
- Генетика
- Диетология
- Дерматовенерология
- Инфекционные болезни
- Культурология
- Лабораторная диагностика
- Летняя практика
- Лучевая диагностика
- Медицинская информатика
- Микробиология
- Неврология
- Общественное здоровье
- Общий уход
О НАС
«Dendrit» - информационный портал для медицинских работников, студентов медицинских ВУЗов, исследователей и пациентов.
Ваш источник новостей и знаний о здоровье.
dendrit.ru © 2014-2024
